Seven Million Years
That is how long this story has been running. At least twenty-one species of human have walked the Earth. We are the only one left.
The Deep Divergence
Somewhere between seven and six million years ago, in the forests of central or eastern Africa, a population of great apes split. One lineage would produce chimpanzees and bonobos. The other would produce us. We do not know what that common ancestor looked like, how it moved, or what it ate. We do not even know, with certainty, where it lived. What we have instead are fragments—a scattering of teeth, a partial cranium, a handful of limb bones—pulled from sediments in Chad, Kenya, and Ethiopia that collectively represent the deepest roots of the hominin family tree.

The oldest candidate is Sahelanthropus tchadensis, known from a single crushed cranium called Toumaï—“hope of life” in the Goran language—unearthed in 2001 from the Djurab Desert of Chad, 2,500 kilometres west of the East African Rift Valley where most paleoanthropologists had expected to find our earliest ancestors. Its foramen magnum, the hole where the spinal cord exits the skull, sits beneath the cranium rather than behind it—a signature of upright posture. Its brain was roughly 350 cubic centimetres, comparable to a modern chimpanzee. The combination is disorienting: bipedalism, it seems, arrived long before the expansion of the brain.
From the Tugen Hills of Kenya came Orrorin tugenensis, dated to roughly six million years ago. Only thirteen fragments survive: pieces of femur, jaw, and finger bones. But the femoral neck shows a cortical bone distribution consistent with habitual bipedalism, and the finger bones curve in a way that suggests tree-climbing had not been abandoned. These creatures lived in a mosaic of forest and open woodland, apparently comfortable in both worlds.
Then, in the Middle Awash of Ethiopia, Tim White and his colleagues recovered Ardipithecus ramidus, a 4.4-million-year-old female nicknamed Ardi. Published in 2009 after fifteen years of painstaking reconstruction, Ardi rewrote expectations. She had an opposable big toe for grasping branches, yet the shape of her pelvis indicated she walked upright on the ground. She was not a knuckle-walker. She was not a creature of the savanna. She lived in closed woodland, eating fruits, nuts, and leaves. The old narrative—that bipedalism evolved on the grasslands, as a response to a drying climate—could no longer hold.

The Australopith Explosion
By four million years ago the hominin lineage had diversified into a radiation of upright-walking primates collectively known as australopiths. They were small—most stood between 1.0 and 1.5 metres tall—and their brains remained modest, hovering around 400 to 500 cubic centimetres. But they had committed to a life on two legs, and that commitment would open every ecological niche that followed.
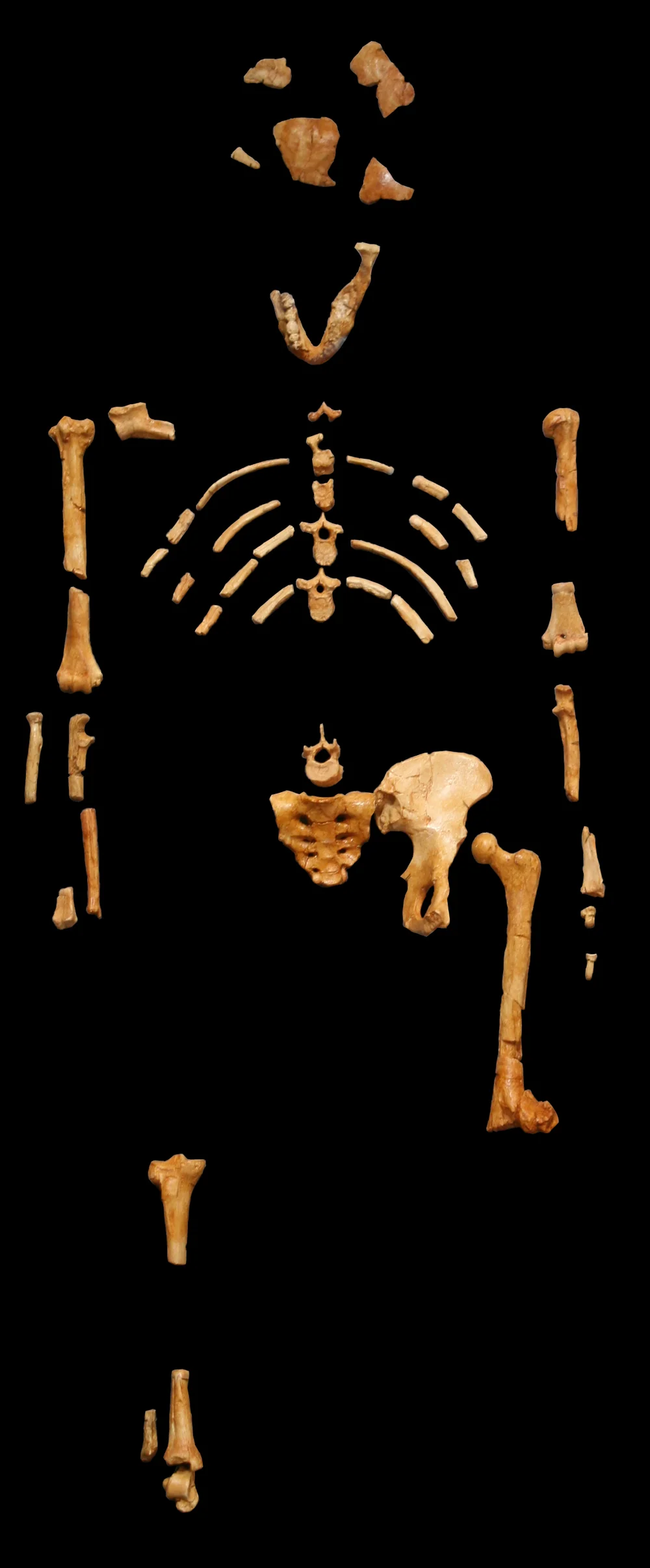
The most celebrated of them is Lucy—AL 288-1—a 3.2-million-year-old partial skeleton of Australopithecus afarensis found at Hadar, Ethiopia, in 1974 by Donald Johanson and Tom Gray. Forty percent of her skeleton survived, an extraordinary proportion for a hominin fossil. Her pelvis was broad and bowl-shaped, her knee angled inward, her gait unmistakably bipedal. The team named her after “Lucy in the Sky with Diamonds,” which was playing at their camp that night. In Amharic she is called Dinkinesh: “you are marvellous.”
A year after Lucy’s discovery, Mary Leakey’s team found something equally stunning 1,500 kilometres to the south: a trail of hominin footprints preserved in volcanic ash at Laetoli, Tanzania, dated to 3.66 million years ago. Two individuals—and possibly a third—had walked side by side across a fresh fall of tuff from the Sadiman volcano. Their prints show a heel strike, a longitudinal arch, and a forward push-off from the big toe. They walked as we walk, more than three million years before the genus Homo would appear.

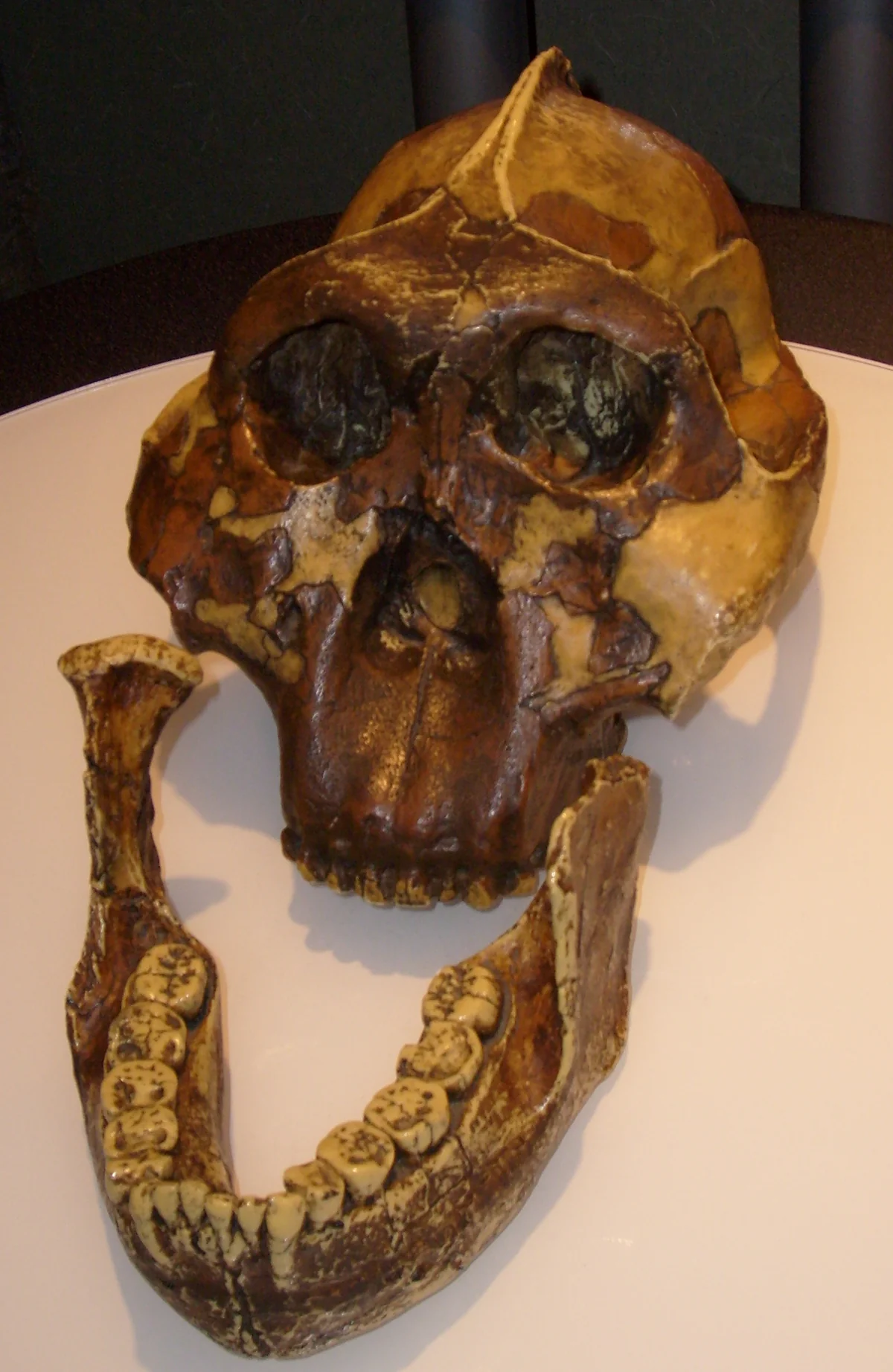
Not all australopiths remained gracile. By 2.7 million years ago a robust lineage had emerged—the genus Paranthropus—characterized by massive jaws, enormous molars, and a sagittal crest anchoring powerful chewing muscles. Paranthropus boisei, nicknamed “Nutcracker Man” when Mary Leakey found the type specimen at Olduvai Gorge in 1959, had a face built for mechanical force. Isotope analysis reveals a diet dominated by C4 plants—grasses and sedges—rather than the hard nuts its morphology seemed designed to crack. Paranthropus would persist for over a million years, coexisting with early Homo, before vanishing without descendants around 1.2 million years ago.
The First Humans
The boundary between australopiths and the genus Homo is one of the most contested lines in paleoanthropology. A fragmentary mandible from Ledi-Geraru, Ethiopia, dated to 2.8 million years ago, represents the oldest known specimen assigned to our genus. Its teeth are smaller and more symmetrical than those of any australopith, yet its jaw still retains primitive proportions. The transition was not a sudden event but a long, messy gradient played out across eastern and southern Africa.
Homo habilis—“handy man”—appeared around 2.4 million years ago and is associated with the Oldowan stone tool industry: simple cobbles struck to produce sharp flakes. Louis Leakey named the species in 1964, convinced that toolmaking was the defining trait of humanity. With a brain averaging 600 cubic centimetres, H. habilis occupied a cognitive no-man’s-land between the australopiths and the larger-brained hominins that followed. Some researchers argue it is not Homo at all but rather a late australopith with a misleading label.
There is no such ambiguity about Homo erectus. Appearing in Africa by 1.9 million years ago, H. erectus was a genuine departure: tall, long-legged, narrow-hipped, with a brain ranging from 600 to over 1,000 cubic centimetres across its 1.5-million-year tenure as a species. The skeleton known as Turkana Boy (KNM-WT 15000), discovered by Kamoya Kimeu at Nariokotome, Kenya, in 1984, died at roughly eight to eleven years of age and already stood 1.6 metres. Had he reached adulthood, he might have exceeded 1.85 metres. His body proportions are essentially modern—built for endurance walking and running in open, tropical landscapes.

And then H. erectus did what no hominin had done before: it left Africa. By 1.8 million years ago, a population had reached Dmanisi in the Republic of Georgia, where five remarkably complete skulls have been recovered from beneath the ruins of a medieval fortress. The Dmanisi hominins were small-brained (546–730 cc) and short, yet they had crossed thousands of kilometres of varied terrain. They carried Oldowan tools, not the more sophisticated Acheulean hand axes that would appear later. The implication is stark: you did not need a large brain or advanced technology to colonise a continent. You needed legs and persistence.
“The question is not why the Neanderthals went extinct. The question is how they managed to survive for 300,000 years.”
The Great Diversification
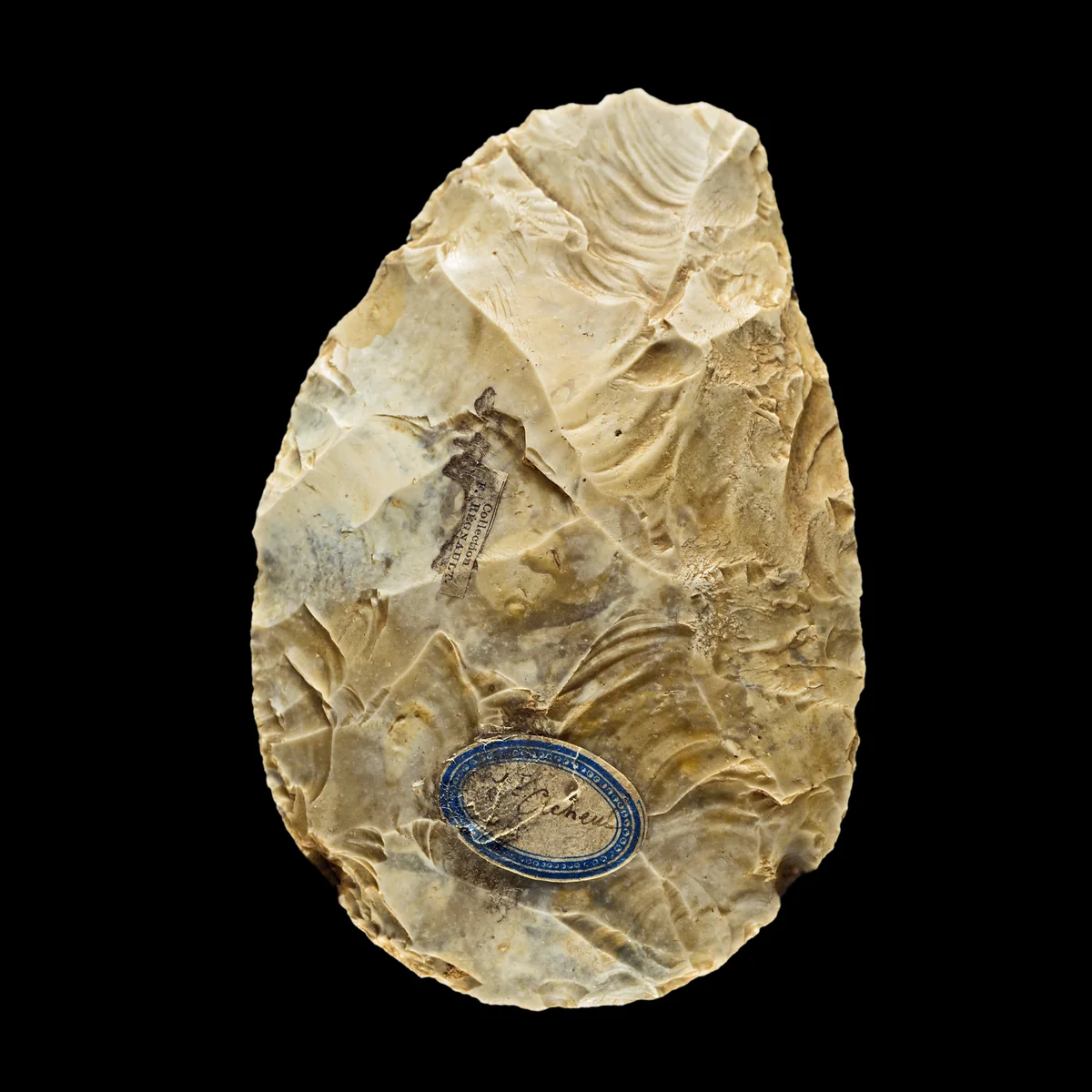
By 800,000 years ago, Homo populations were scattered across Africa, Europe, and much of Asia—and they were diverging. The Middle Pleistocene is the most tangled chapter of hominin evolution, a period when at least five distinct lineages occupied overlapping ranges and the fossil record offers just enough evidence to provoke argument but not enough to settle it. At the centre of this radiation stands Homo heidelbergensis, a species defined by the Mauer mandible found near Heidelberg, Germany, in 1907. With brains ranging from 1,100 to 1,400 cubic centimetres and robust builds adapted to cold climates, these hominins hunted large game cooperatively, built shelters, and controlled fire with increasing sophistication.
At the site of Schöningen in northern Germany, archaeologists recovered eight wooden spears dated to approximately 300,000 years ago—the oldest known throwing weapons. Each was carved from a single spruce trunk, with the centre of gravity placed one-third of the way from the tip, exactly as in modern javelins. These were not opportunistic stakes. They were engineered projectiles, products of foresight and accumulated technical knowledge. Their makers killed horses at a lakeside ambush and butchered them on the spot. The site preserves no symbolic artefacts, no ochre, no beads—but the planning depth is unmistakable.
Fire transformed everything. Evidence from Wonderwerk Cave in South Africa pushes controlled combustion back to at least one million years ago, but by the Middle Pleistocene fire use was habitual and widespread. Cooking softened food, unlocking more calories from the same intake and reducing the metabolic cost of digestion. Richard Wrangham’s “cooking hypothesis” argues that this caloric surplus was the primary driver of brain expansion in Homo—not toolmaking, not sociality, but fire and the kitchen it created. The archaeological record does not yet confirm every link in that chain, but the correlation between habitual fire use and accelerating encephalization is difficult to dismiss.
By the end of this period, multiple hominin species occupied the Old World simultaneously. In Africa, archaic populations that would eventually give rise to Homo sapiens were evolving a suite of modern skeletal features. In Europe, cold-adapted populations were becoming Neanderthals. In eastern Asia, the mysterious Denisovans were leaving their genetic signature in mountain caves. On the island of Flores, a diminutive hominin with a 380-cc brain was crafting stone tools and hunting pygmy elephants. The tree of human evolution was not a ladder. It was a bramble.
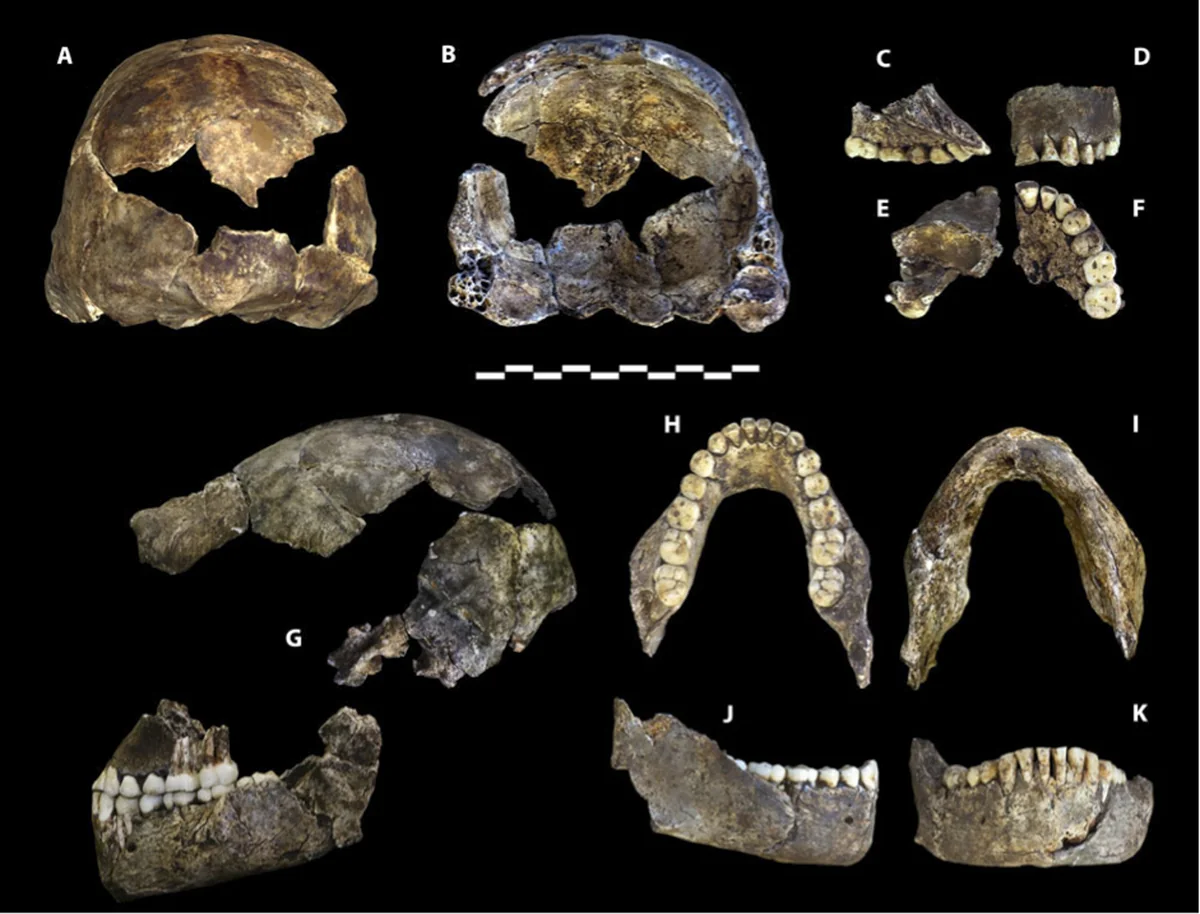
Our Closest Kin
The Neanderthals
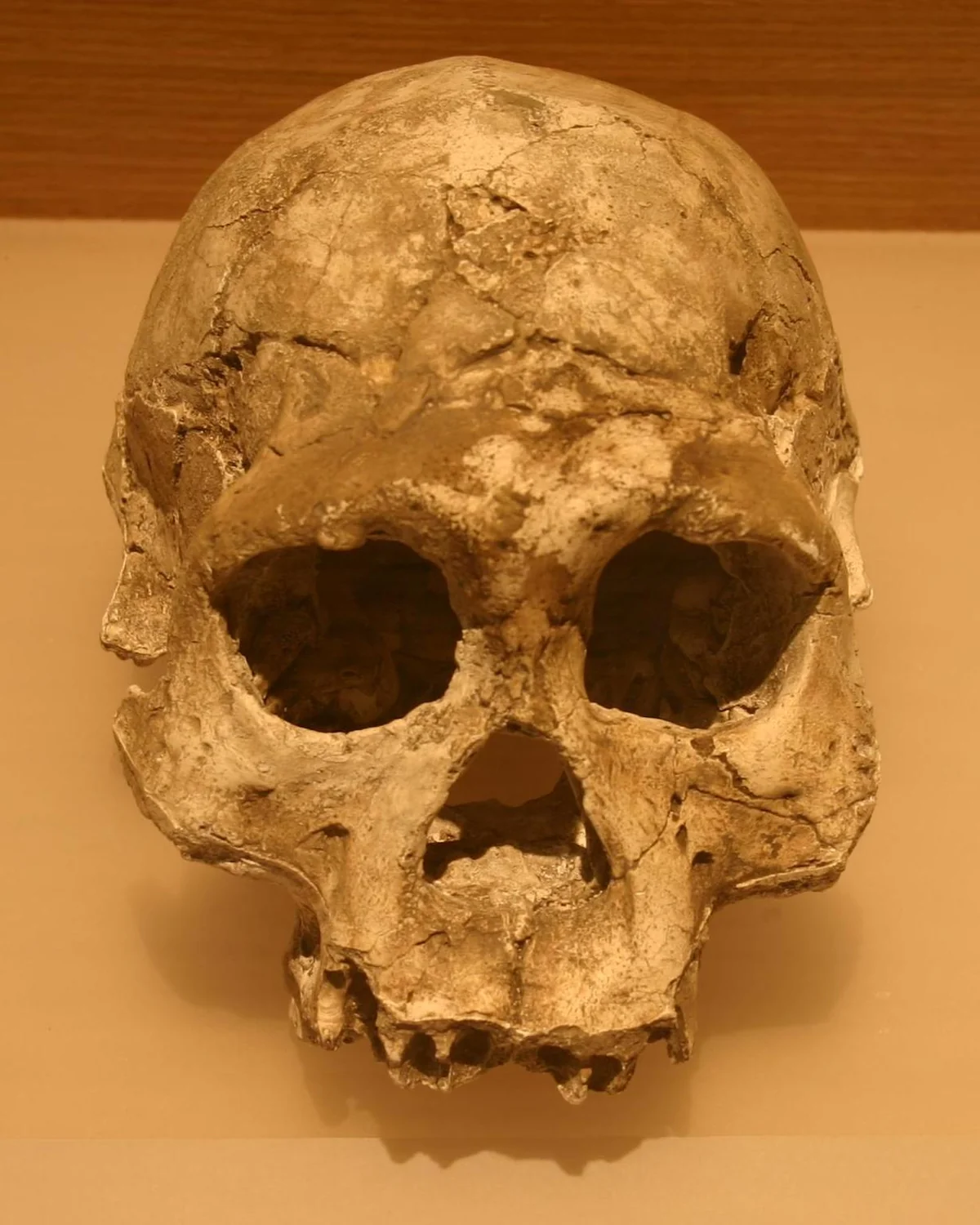
No extinct hominin has been as thoroughly studied—or as persistently caricatured—as Homo neanderthalensis. For a century after their recognition in the 1860s, Neanderthals were depicted as slouching, dim-witted brutes: the evolutionary dead end that Homo sapiens had rightly replaced. That image was wrong in virtually every particular. Neanderthals had brains averaging 1,450 cubic centimetres—larger than our own. They were powerfully built, barrel-chested, with bones so thick their cross-sections resemble those of Olympic weight-lifters. They endured Ice Age Europe for more than 300,000 years, tracking mammoths and woolly rhinoceroses through boreal forests and tundra in conditions that would kill an unequipped modern human in hours.
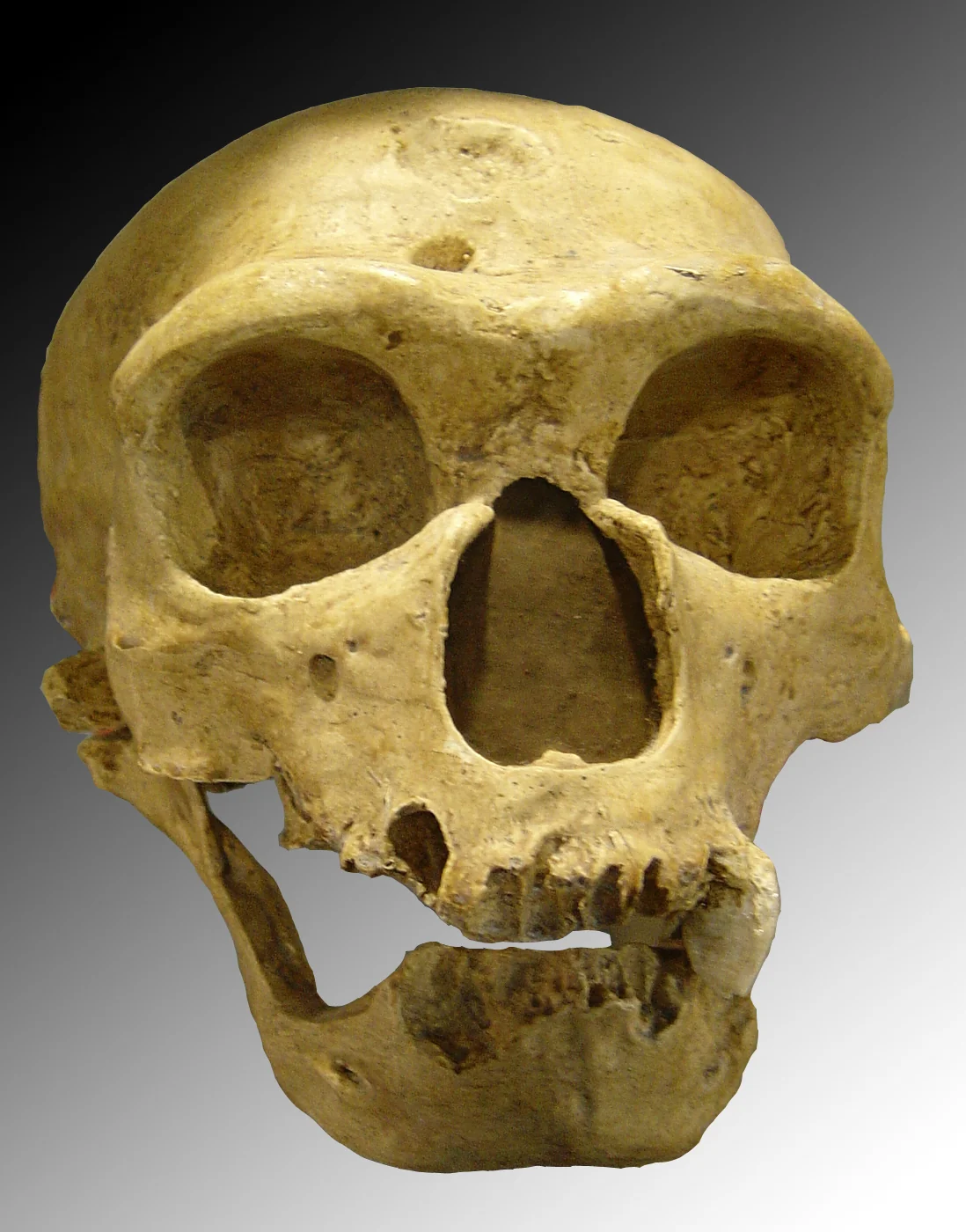
Their technology was sophisticated. The Levallois technique—a prepared-core method of flint-knapping that requires the knapper to hold a three-dimensional mental template of the desired flake before striking—dominated Neanderthal assemblages across Europe and western Asia. They hafted stone points onto wooden shafts using birch-bark pitch, a thermoplastic adhesive whose manufacture demands precise temperature control (340–400 °C) in an oxygen-reduced environment. Experimental archaeologists have struggled to replicate the process reliably.

They used ochre pigments at sites like Maastricht-Belvdère. At Bruniquel Cave in southwestern France, deep inside a chamber 336 metres from the entrance, they constructed ring-shaped structures from broken stalagmites 176,000 years ago—the oldest known deliberate constructions in the subterranean world.
They also buried their dead. At Shanidar Cave in Iraqi Kurdistan, Ralph Solecki excavated Neanderthal skeletons in the 1950s and 1960s, one of which appeared to be surrounded by flower pollen—a finding that spawned the romantic image of the “flower burial.” More recent excavations at the same site by Graeme Barker’s team have confirmed deliberate burial but cast doubt on the flowers. What is not in doubt is that Neanderthals cared for their injured and infirm. The Shanidar 1 individual had survived severe crushing injuries to the left side of his skull, withering of his right arm, and damage to both legs—injuries that would have required sustained assistance from others over months or years.
And yet they disappeared. Between 45,000 and 40,000 years ago, Neanderthal populations across Europe contracted and vanished, a process that took roughly five millennia—an eye-blink in geological time but several hundred generations of lived experience. Their final refugia appear to have been the Iberian Peninsula and possibly the southern Balkans. The exact mechanism of their extinction remains one of the most intensely debated questions in palaeoanthropology. Climate instability, competition with incoming sapiens populations, demographic fragility in small groups, disease transfer—all have been proposed, and none alone suffices.
What the genomic evidence has made clear is that Neanderthals did not simply vanish. They were partially absorbed. Between 1 and 4 percent of the genome of every living non-African human derives from Neanderthal ancestors, the product of interbreeding events that occurred when sapiens populations moved into western Asia and Europe. Neanderthal alleles persist in us today, influencing traits from immune function to skin and hair pigmentation to the shape of our skulls. They are gone as a species, but they endure as a ghostly fraction of our biology.
The Rise of Sapiens
In 2017, Jean-Jacques Hublin and his team published a redating of fossils from Jebel Irhoud, a collapsed cave in the hills west of Marrakech, Morocco. The skulls, originally excavated in the 1960s and dismissed as anomalous Neanderthals, turned out to be 315,000 years old—the earliest known Homo sapiens. Their faces were flat and retracted, essentially modern; their braincases were elongated and archaic. The find pushed the origin of our species back by 100,000 years and, critically, placed it in North Africa rather than East Africa. Combined with similarly archaic-modern mosaics from Florisbad in South Africa (259 kya) and Omo Kibish in Ethiopia (233 kya), the picture that emerged was of a pan-African origin: sapiens did not evolve in a single Eden but across the continent, in populations connected by intermittent gene flow across a patchwork of habitable corridors.
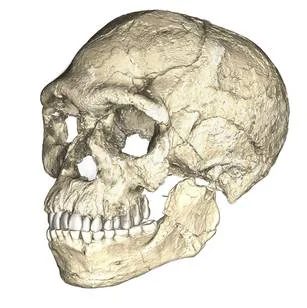
The earliest sapiens were anatomically transitional but not yet behaviourally modern in the archaeological sense. Perforated shell beads appear at Blombos Cave in South Africa by 75,000 years ago; ochre crayons with geometric cross-hatchings are older still, dated to 100,000 years. In the Kalahari, crystal collections and pigment processing date to 105,000 years ago at Ga-Mohana Hill. Symbolic thought—the capacity to let one thing stand for another—was not a sudden mutation. It accumulated in fragments over tens of thousands of years, expressed differently in different populations, flickering in and out of the archaeological record like a signal that only slowly resolved into something continuous.

By 70,000 years ago, sapiens populations in southern Africa were manufacturing backed microliths—tiny stone blades intended to be hafted in composite tools—at Pinnacle Point and Sibudu Cave. The technology required planning across multiple stages: selecting raw material, heat-treating silcrete to improve its flaking properties, knapping the blanks, shaping the elements, preparing mastic adhesive, and assembling the finished implement. Each step drew on transmitted knowledge. Alone, none of these operations is beyond Neanderthal capability. Chained together, they represent a density of cultural transmission that appears to have no parallel in the Neanderthal record.
The Coexistence
For most of hominin history, multiple species shared the planet. But the period between 130,000 and 40,000 years ago represents the last great age of coexistence, a stretch during which at least five distinct lineages overlapped in time and, in some cases, in space. Homo sapiens occupied Africa and the Levant. Homo neanderthalensis ranged across Europe and western Asia. The Denisovans, known almost entirely from DNA extracted from finger bones and teeth in a Siberian cave, inhabited a vast arc from the Altai Mountains to Southeast Asia. Homo floresiensis, the “Hobbit,” persisted on the island of Flores until at least 60,000 years ago. And Homo naledi, with its small brain and surprisingly modern hands and feet, was depositing its dead in the Rising Star cave system of South Africa as recently as 236,000 years ago.

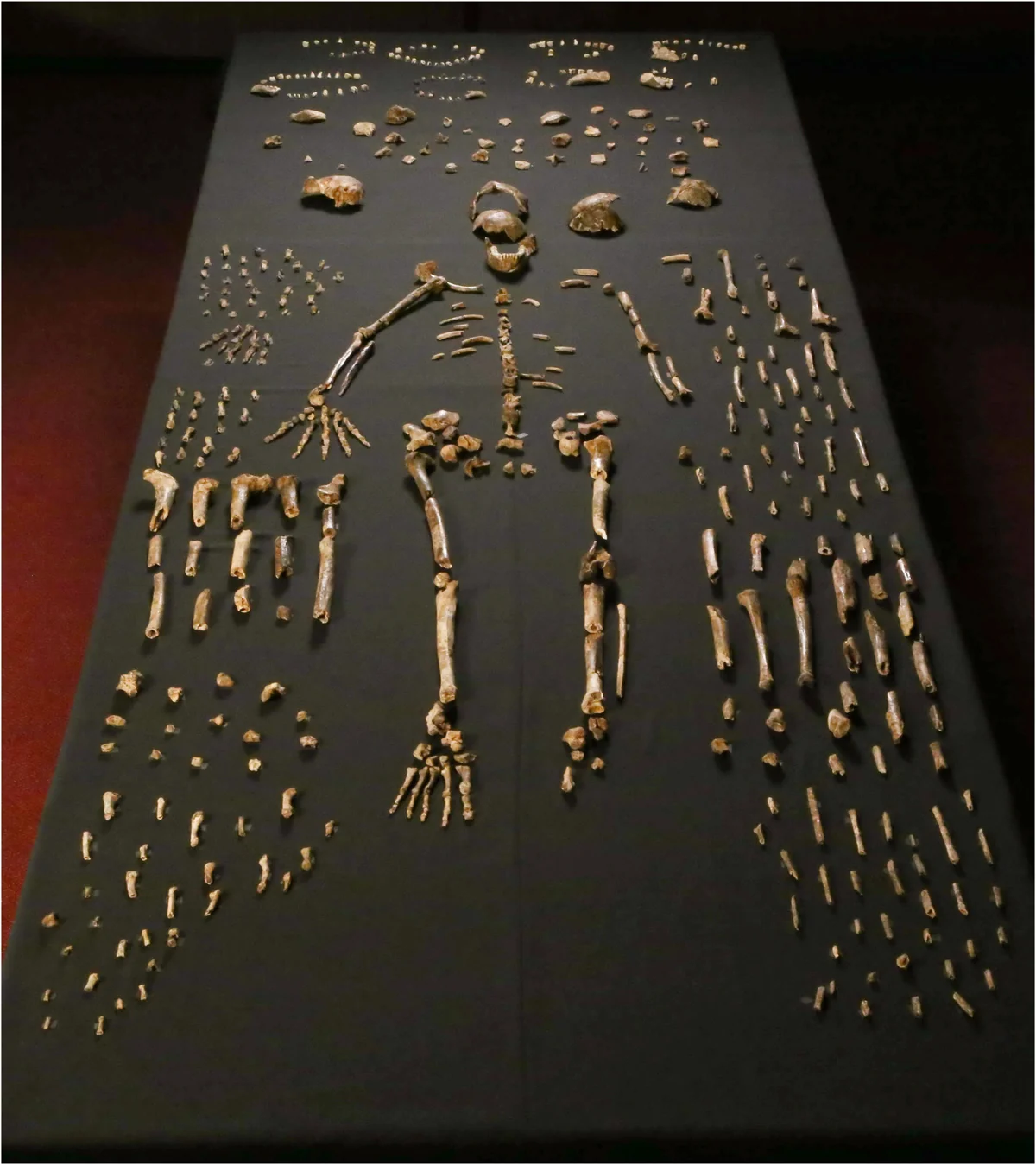
These species did not merely coexist. They interbred. The sequencing of ancient DNA—pioneered by Svante Pääbo and his team at the Max Planck Institute in Leipzig, work that earned the 2022 Nobel Prize in Physiology or Medicine—revealed gene flow between sapiens and Neanderthals, between sapiens and Denisovans, and between Denisovans and Neanderthals. A single bone fragment from Denisova Cave (Denisova 11) belonged to a first-generation hybrid: a girl whose mother was Neanderthal and whose father was Denisovan. The old metaphor of a family tree fails entirely. A braided stream is closer to the truth.
Why We Survived
Of more than twenty hominin species that have existed since the divergence from our last common ancestor with chimpanzees, exactly one persists. The question of why Homo sapiens alone survived is not settled, and any honest answer must accommodate uncertainty. But decades of research across genetics, archaeology, ecology, and climate science have converged on a set of factors that, taken together, provide a plausible framework.
First, social networks. Sapiens maintained larger and more interconnected social groups than any contemporary hominin. Neanderthal groups rarely exceeded 15–25 individuals, based on site-occupation signatures and genetic diversity estimates. Sapiens bands were comparable in size but were embedded in wider networks of exchange and alliance that linked hundreds or thousands of individuals across regions. These networks transmitted information, buffered local resource failures, and provided mating partners who prevented the inbreeding depression that plague small, isolated populations.
Second, language—though its role is the most difficult to assess from the archaeological record. Neanderthals possessed a hyoid bone consistent with speech and shared with sapiens the FOXP2 gene variant associated with language capacity. Whether their communication was fully syntactic or more limited remains unknown. What the demographic and technological record suggests is that sapiens were better at encoding, storing, and transmitting complex procedural knowledge across generations—a capability that depends on the precision and flexibility of language.
Third, dietary flexibility. Isotope analyses reveal that Neanderthals were apex predators whose diets were dominated by large herbivores—mammoth, bison, horse, deer. Sapiens exploited a broader spectrum: shellfish, fish, birds, small mammals, seeds, tubers, insects. When large-game populations collapsed during climatic oscillations, Neanderthal populations starved. Sapiens switched menus. Fourth, the ratchet of cultural cumulation described above: each sapiens generation reliably inherited and built upon the innovations of the last. Fifth, demographic advantage—sapiens populations in Africa were larger, more genetically diverse, and better buffered against extinction. And sixth, an element that no theory can fully account for: luck. Volcanic winters, disease events, and stochastic population crashes likely eliminated hominin groups irrespective of their adaptations. Survival at the species level is not always a verdict on fitness. Sometimes it is a coin flip.
The Migration
Homo sapiens left Africa multiple times. An early dispersal reached the Levant by 177,000 years ago (Misliya Cave, Israel), but that population apparently did not persist. The sustained expansion began around 70,000–60,000 years ago, likely via the southern route across the Bab el-Mandeb strait into the Arabian Peninsula. From there, the colonization of the globe proceeded with astonishing speed. Within 50,000 years—a fraction of a percent of total hominin history—sapiens had reached every major landmass except Antarctica.
The feat demanded maritime technology (to reach Australia and the Pacific islands), cold-weather adaptation (to survive Arctic Siberia), and the cognitive capacity to inhabit radically different ecosystems within a few thousand years. No other primate has accomplished anything comparable. No other hominin came close.
“We are walking archives of ancestral wisdom.”
The Last Ones Standing
For approximately 40,000 years, Homo sapiens has been the sole surviving member of a lineage that once numbered at least twenty-one species. We are accustomed to this solitude—it is the only condition any living human has known—but in the deep context of hominin history, it is the exception. For most of the seven million years covered by this essay, multiple human species shared the planet. They occupied different niches, adapted to different climates, invented different tools, and in some cases recognized one another well enough to interbreed. The world they inhabited was richer in human diversity than anything we can experience today. Their extinction impoverished the Earth in ways we cannot fully measure.
We carry their legacy in our genomes. Every person of non-African descent bears Neanderthal alleles that influence the structure of their immune system, the keratin in their hair, and their susceptibility to certain diseases. Melanesian and Aboriginal Australian populations carry Denisovan variants that help them metabolize oxygen at high altitude and resist tropical pathogens. African populations, long assumed to have no archaic admixture, have been shown to carry fragments of DNA from at least one unknown hominin lineage that diverged from our own more than 500,000 years ago. We are not purely sapiens. We are composites—braided streams of genetic information drawn from branches of the human family that no longer exist as independent species. The dead walk with us in every cell.
Sources & Further Reading
- 1.Brunet, M., et al. (2002). A new hominid from the Upper Miocene of Chad, Central Africa. Nature, 418(6894), 145–151.
- 2.White, T.D., et al. (2009). Ardipithecus ramidus and the Paleobiology of Early Hominids. Science, 326(5949), 64–86.
- 3.Johanson, D.C. & White, T.D. (1979). A systematic assessment of early African hominids. Science, 203(4378), 321–330.
- 4.Leakey, M.G., et al. (2001). New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature, 410(6827), 433–440.
- 5.Berger, L.R., et al. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife, 4, e09560.
- 6.Brown, P., et al. (2004). A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature, 431(7012), 1055–1061.
- 7.Hublin, J.-J., et al. (2017). New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature, 546(7657), 289–292.
- 8.Green, R.E., et al. (2010). A Draft Sequence of the Neandertal Genome. Science, 328(5979), 710–722.
- 9.Reich, D., et al. (2010). Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature, 468(7327), 1053–1060.
- 10.Stringer, C. (2012). Lone Survivors: How We Came to Be the Only Humans on Earth. Times Books.
- 11.Wood, B. & Richmond, B.G. (2000). Human evolution: taxonomy and paleobiology. Journal of Anatomy, 197(Pt 1), 19–60.
- 12.Antón, S.C., Potts, R. & Aiello, L.C. (2014). Evolution of early Homo: An integrated biological perspective. Science, 345(6192), 1236828.
- 13.Tattersall, I. (2012). Masters of the Planet: The Search for Our Human Origins. Palgrave Macmillan.
- 14.Potts, R. (1998). Variability Selection in Hominid Evolution. Evolutionary Anthropology, 7(3), 81–96.
- 15.Lordkipanidze, D., et al. (2013). A Complete Skull from Dmanisi, Georgia, and the Evolutionary Biology of Early Homo. Science, 342(6156), 326–331.
- 16.Détroit, F., et al. (2019). A new species of Homo from the Late Pleistocene of the Philippines. Nature, 568(7751), 181–186.
- 17.Ni, X., et al. (2021). Massive cranium from Harbin in northeastern China establishes a new Middle Pleistocene human lineage. The Innovation, 2(3), 100130.
- 18.Harmand, S., et al. (2015). 3.3-million-year-old stone tools from Lomekwi 3, West Turkana, Kenya. Nature, 521(7552), 310–315.
- 19.Prüfer, K., et al. (2014). The complete genome sequence of a Neanderthal from the Altai Mountains. Nature, 505(7481), 43–49.
- 20.Rightmire, G.P. (2004). Brain size and encephalization in early to Mid-Pleistocene Homo. Am. J. Phys. Anthropol., 124(2), 109–123.
- 21.Mellars, P. (2006). Why did modern human populations disperse from Africa ca. 60,000 years ago? PNAS, 103(25), 9381–9386.
- 22.Berger, L.R., et al. (2010). Australopithecus sediba: A New Species of Homo-Like Australopith from South Africa. Science, 328(5975), 195–204.
- 23.Dunbar, R.I.M. (1998). The social brain hypothesis. Evolutionary Anthropology, 6(5), 178–190.
Image Credits
- Sahelanthropus tchadensis cranium — Didier Descouens, CC BY-SA 4.0
- Ardipithecus ramidus skeleton cast — Tiia Monto, CC BY-SA 4.0
- Lucy skeleton (AL 288-1) — 120, CC BY-SA 3.0
- Laetoli footprints replica — Tim Evanson, CC BY-SA 2.0
- Paranthropus boisei skull — Durova, CC BY-SA 4.0
- Olduvai Gorge panorama — Noel Feans, CC BY 2.0
- Turkana Boy skeleton — Bone Clones, CC BY-SA 3.0
- Acheulean hand axe — Didier Descouens, CC BY-SA 4.0
- Homo naledi skeletal specimens — Lee Roger Berger et al., CC BY 4.0
- Neanderthal cranium — Luna04, CC BY-SA 2.0
- Homo floresiensis skull — Ryan Somma, CC BY-SA 2.0
- Lion Man of Hohlenstein-Stadel — Dagmar Hollmann, CC BY-SA 4.0
- Dmanisi skull (1.8 MYA) — Gerbil, CC BY-SA 4.0
- Levallois flake from Nesher Ramla — Tal Rogovski, CC BY-SA 4.0
- Homo sapiens from Jebel Irhoud — NHM London, CC BY-SA 4.0
- Homo naledi holotype cranium (DH1) — Lee Roger Berger et al., CC BY 4.0
- Great Rift Valley aerial view — Wikimedia Commons, CC BY 2.0
All images sourced from Wikimedia Commons under Creative Commons or Public Domain licenses. Cranial capacity and body size data drawn from Holloway et al. (2004), Grabowski et al. (2015), and Antón et al. (2014). Comparative trait scores are normalized estimates adapted from the frameworks of Finlayson (2004), Stringer (2012), and Wynn & Coolidge (2012).