
Turkana Boy: The First Modern Body
108 Bones, a Hole in the Jaw, and the Child Who Carried Our Body Plan Into the Open World
The Abscess
“You can read a skeleton the way a coroner reads a body.”
Below the left first molar of specimen KNM-WT 15000, there is a hole in the jawbone. It is roughly the size of a fingertip — a cavity of remodelled bone surrounding the root apex, where infection had eaten through the mandibular cortex and established a draining sinus. The bone around it is thickened and irregular, the telltale signature of chronic periapical abscess: the body had mounted an inflammatory response, laid down new bone to wall off the infection, and sustained that defense long enough for the reaction to become architecturally visible 1.53 million years later.
This is where the story begins — not with a species or an epoch, but with a lesion. Alan Walker, who led the anatomical analysis of the skeleton across nearly a decade of study, described the infection as severe and chronic. The remodelled bone indicated weeks or months of active disease. Whether the abscess contributed to death is debatable; Walker himself acknowledged that the causal chain from infection to mortality cannot be established from bone alone. What the lesion does establish, with descriptive certainty, is that this individual was in pain. The left side of the mandible carried the evidence of sustained suffering — a biological fact readable from the morphology of the bone itself, independent of any interpretive framework.
The distinction matters. Paleopathology — the study of disease in ancient remains — operates within a strict hierarchy of inference. At the base sits description: the hole is there, the bone is remodelled, the dimensions are measurable. One step up sits diagnosis: the pattern is consistent with periapical abscess secondary to dental infection. Further up, and the ground becomes less certain: was this the cause of death? Did sepsis spread from the jaw to the bloodstream? Did the infection compromise his ability to eat, weakening him over weeks? These are plausible inferences, not demonstrated facts. Brown and colleagues, in their original 1985 description inNature, reported the lesion alongside the specimen’s geological context and taxonomic assignment but stopped short of causal claims about mortality. Walker, writing in the 1993 monograph, went further in characterizing the infection’s severity but maintained the same epistemic boundary: we can describe what we see; we cannot narrate what we cannot observe.
Beginning with the abscess is a deliberate narrative choice. Most accounts of Turkana Boy open with triumph — the most complete early Homo skeleton ever found, 108 bones from a single individual, a specimen that transformed paleoanthropology. That framing is accurate but incomplete. It places the skeleton in the story of science rather than in the story of a life. The abscess reverses the emphasis. Before this skeleton was a dataset, it was a child with an infected jaw. Before it answered questions about human evolution, it carried the evidence of an individual experience — pain, inflammation, the slow biological response of bone to bacterial invasion.
The essay that follows will move from this forensic starting point outward through anatomy, development, ecology, cognition, and dispersal. But the method established here — describe first, interpret carefully, acknowledge what remains unknown — will govern every section. When we reach questions about adult stature, growth rate, speech capacity, or locomotor efficiency, the same hierarchy applies. The skeleton tells us what it tells us. Our job is to read it honestly, not to make it say more than it can.
Skeletal Completeness by Region
Data: Walker & Leakey 1993; Brown et al. 1985“The skeleton was unlike anything we had seen from that time period. Not just in preservation — in completeness. We had a cranium, a mandible, vertebrae, ribs, pelvis, limb bones. For the first time we had a whole person.”
— Frank Brown, Nature, 1985
The Age
“The teeth say one thing, the bones say another.”
How old was he when he died? The answer depends on which clock you read. The skeleton of KNM-WT 15000 carries two independent chronometers — dental development and skeletal maturation — and they disagree by roughly three years. That discrepancy is not a footnote. It is the intellectual fulcrum on which nearly every downstream claim about this specimen pivots: projected adult height, body mass, brain-to-body ratio, metabolic requirements, and the growth schedule of his entire species.
Holly Smith, in her 1993 analysis of the specimen’s dentition, established the framework that still governs the debate. The second molars were in the process of erupting. The third molars had not yet begun their descent. By modern human standards, this pattern corresponds to an age of approximately eleven years. But Smith recognized that modern human developmental timing might be the wrong yardstick. If earlyHomo ergaster matured on a schedule closer to the great apes — as growing evidence from dental microstructure suggested — then the same eruption pattern would correspond to an age of approximately eight years, perhaps slightly less. The same teeth, read against different calibration standards, produced two fundamentally different ages.
The skeletal evidence, analyzed by Christopher Ruff and Alan Walker, pointed toward the older estimate. Long bone lengths, when compared to modern human growth charts, suggested a boy of eleven to twelve. Epiphyseal fusion status — the degree to which the growth plates at the ends of the bones had closed — was consistent with a pre-adolescent on a modern human timeline. But Ruff and Walker were explicit about the circularity: if the species matured faster than modern humans, then the same bone lengths and fusion status could belong to a younger individual who was simply larger for his age.
In 2009, M. Christopher Dean and Holly Smith revisited the question using dental histology — the microscopic growth lines preserved in tooth enamel called perikymata. Each perikymata line represents approximately one week of enamel deposition, providing a direct biological clock independent of comparative eruption schedules. Their analysis indicated that dental formation in KNM-WT 15000 proceeded significantly faster than in modern humans, supporting an age at death closer to eight years. The developmental pattern was intermediate between apes and humans but closer to the ape end of the spectrum. This was not simply a matter of choosing between two ages. It was a finding about the species itself: the prolonged childhood that we consider essentially human had not yet evolved by 1.5 million years ago.
The consequences cascade. If he was eight, he was near the end of his growth — a large juvenile approaching adult size on a fast maturation schedule. If he was eleven, he had years of growth remaining and would have become extraordinarily tall. Every estimate of adult stature, body mass, encephalization quotient, and energetic budget depends on which age you adopt. The skeleton does not contain a single individual; it contains a developmental argument. And that argument remains genuinely unresolved.

The Age Dispute: Dental vs. Skeletal Estimates
Data: Smith 1993; Dean & Smith 2009; Walker & Leakey 1993; Ruff & Walker 1993“The teeth say one thing, the bones say another. If we age him by dental eruption using modern human standards, he was about eleven. If we calibrate for faster development — closer to the great-ape pattern — he was eight, perhaps nine. The difference is not trivial. It changes the entire reconstruction.”
— Holly Smith, in Walker & Leakey (eds.), 1993
The Growth
“At 160 centimeters he was already taller than many adult australopiths. But his skeleton was unfinished.”
At approximately 160 centimeters — five feet three inches — KNM-WT 15000 was already taller at death than any adult Australopithecus afarensis ever measured, taller than the reconstructed stature of most Homo habilis specimens, and within the range of adult stature estimates for the Dmanisi Homo erectus population that had already dispersed to the Caucasus. He was a child who had outgrown adults of earlier species. And he was not finished growing.
The unfused epiphyses — the growth plates at the ends of his long bones — confirm that active growth was still underway at death. The question is how much growth remained, and the answer depends entirely on the age model adopted in the previous section. Christopher Ruff and Alan Walker, using modern human growth curves and the older skeletal age estimate of eleven to twelve years, projected an adult stature of approximately 185 centimeters — over six feet tall. Under this slow-maturation model, Turkana Boy still had an adolescent growth spurt ahead of him, and his projected adult body mass would have reached 62 to 68 kilograms.
The fast-maturation model tells a different story. If dental histology is correct and he was approximately eight years old, then his growth curve was far steeper than a modern child’s, and he was already approaching the asymptote. Dean and Smith’s analysis, combined with Ruff and Burgess’s 2015 reassessment, suggests that under rapid maturation the projected adult stature falls to roughly 163 to 170 centimeters — tall by Pleistocene standards, but not the giant implied by the slow model. An intermediate model, splitting the difference, yields something around 175 to 180 centimeters.
The uncertainty is not a failure of the evidence. It is the evidence. The skeleton captures a moment in a growth trajectory whose tempo was neither fully human nor fully ape-like, but something intermediate — a developmental schedule that had begun to decelerate relative to great apes but had not yet acquired the prolonged adolescence characteristic of modern humans. That intermediate growth tempo is itself one of the most important findings about Homo ergaster. It tells us that the evolution of human childhood — the long, slow developmental period that allows extended learning and neural maturation — was still in progress at 1.53 million years ago.
Projected Growth Trajectories
Data: Ruff & Walker 1993; Dean & Smith 2009; Ruff & Burgess 2015; WHO 2006“If his growth followed the modern human pattern, he was an eleven-year-old boy who would have been over six feet tall. If he grew like an ape, he was eight and already close to his adult size. The answer changes everything downstream — adult body mass estimates, brain-to-body ratios, energetics.”
— Christopher Ruff & Alan Walker, 1993
The Body
“This body dissipates heat like a radiator.”
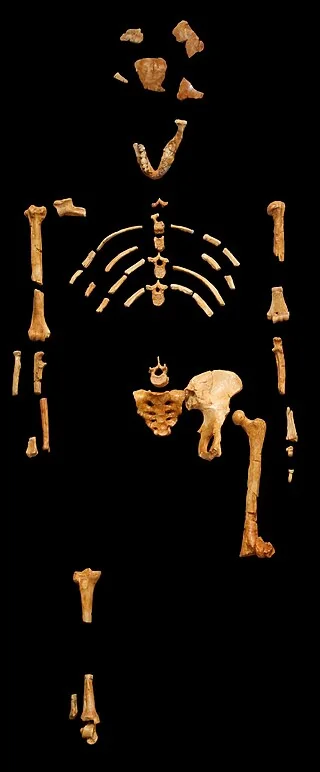
Place Lucy’s skeleton next to Turkana Boy’s and you are not simply looking at a bigger version of the same animal. You are looking at a different engineering solution to a different ecological problem. Australopithecus afarensis, represented most famously by the 3.18-million-year-old partial skeleton AL 288-1, stood roughly 107 centimeters tall, weighed approximately 29 kilograms, and carried a body built for compromise — short legs still suited for arboreal climbing, wide flaring ilia providing attachment surfaces for muscles that stabilized a pelvis not yet fully committed to the mechanical demands of habitual bipedalism. Her bi-iliac breadth relative to stature was approximately 0.25 — a wide, compact body that retained heat efficiently and maximized stability in a mixed woodland-savanna environment.
KNM-WT 15000 is a different architecture. Even at his juvenile stature of 160 centimeters, his proportions had shifted categorically. His bi-iliac breadth — the width across the hip bones — was approximately 23 centimeters, giving a breadth-to-stature ratio of roughly 0.14. This is dramatically narrower than any australopith, narrower than the average modern human ratio of 0.16, and comparable to the proportions seen today in the tallest, most linear tropical populations such as the Nilotic peoples of South Sudan. His crural index — the ratio of tibial length to femoral length, a reliable indicator of limb segment proportionality and climatic adaptation — was approximately 86, squarely within the range of modern equatorial Africans and well above the values recorded for A. afarensis.
Peter Wheeler, in a pair of landmark papers published in 1991 and 1993, provided the mechanistic explanation for why these proportions matter. A tall, narrow body standing upright in equatorial sun presents a minimal cross-section to overhead solar radiation. The high surface-area-to-mass ratio maximizes convective heat loss — the body sheds heat more efficiently because there is proportionally more skin surface through which excess thermal energy can escape. Wheeler calculated that a body built like Turkana Boy’s would gain approximately 30% less solar heat and require roughly 30% less water to maintain thermal balance compared to a shorter, wider body of equivalent mass under the same equatorial conditions. In a hot, dry, open landscape — precisely the habitat that the Turkana Basin represented at 1.53 million years ago — that is not a marginal advantage. It is the difference between sustainable ranging and heat death.
This is Allen’s Rule made visible in the fossil record. First articulated in 1877, Allen’s Rule observes that endothermic animals in hot climates tend toward longer extremities and more linear body forms, while those in cold climates tend toward shorter limbs and more compact bodies — maximizing heat dissipation in the first case, minimizing it in the second. The rule describes a pattern observed across dozens of mammalian species, from foxes to rabbits to humans. In modern Homo sapiens, the pattern is clearly visible: Nilotic East Africans have the highest crural indices and most linear body forms, while Arctic-adapted populations like the Inuit have the lowest crural indices and most compact builds. Turkana Boy represents the first appearance of this heat-adapted body plan in the hominin fossil record — the moment when our lineage abandoned the arboreal-terrestrial compromise body and committed to an open-country thermoregulatory design.
Body Proportion Comparison
Data: Ruff & Walker 1993; Jungers 1988; WHO 2006Thermoregulation: Surface-to-Mass Ratio & Heat Dissipation
Data: Wheeler 1991, 1993; Ruff 1991“When you compare his skeleton to Lucy’s, you are not just seeing a bigger animal. You are seeing a different solution to a different problem. Lucy’s body was a compromise between the ground and the trees. This boy’s body has resolved the compromise. The trees are gone from his anatomy.”
— Alan Walker, The Nariokotome Homo erectus Skeleton, 1993
The Legs
“When you look at those femora, you stop thinking about walking and start thinking about ranging.”
Lucy walked. There is no serious debate about that — the Laetoli footprints, the valgus angle of her knee, the anterior position of her foramen magnum all confirm habitual bipedal locomotion in Australopithecus afarensis by at least 3.6 million years ago. But Lucy walked with a body that retained significant arboreal compromises: relatively short legs, curved phalanges on both hands and feet, a wide pelvis that required pronounced lateral pelvic tilt during swing phase. Her bipedalism was real, but it was expensive. The cost of transport — the metabolic energy required to move one kilogram of body mass over one meter of ground — was substantially higher than in modern humans, perhaps 50% higher by some estimates.
Turkana Boy’s legs tell a different story. His femoral length of approximately 432 millimeters at death was already within 5% of the average adult modern human male value of 455 millimeters — remarkable given that he was a juvenile with unfused epiphyses. His tibia was proportionally long, yielding a crural index of approximately 86, indicating that the distal limb segment had elongated relative to the proximal segment in exactly the pattern predicted by Allen’s Rule for a heat-adapted, open-country biped. His femoral head was 41 millimeters in diameter, reflecting the increased joint loading associated with a larger, heavier body traveling farther distances. His bicondylar angle — the angle of the femoral shaft relative to the condyles at the knee — was approximately 11 degrees, within the modern human range and indicating fully committed valgus alignment for efficient bipedal weight transfer.

Herman Pontzer, in his 2012 analysis of ecological energetics in early Homo, quantified the implications. Longer legs and a more efficient gait reduced the cost of transport in Homo ergaster/erectus by roughly 50% compared to australopiths. A body like Turkana Boy’s could cover the same distance as Lucy’s at half the metabolic cost — or, more relevantly, could cover twice the distance for the same caloric expenditure. This is not an abstract biomechanical calculation. It is the energetic foundation of a new ecological strategy. Australopiths were local foragers, tied to relatively small home ranges in mosaic woodland-savanna environments. A body with Turkana Boy’s locomotor efficiency could range widely across open landscapes, accessing dispersed food resources — including animal carcasses — that would have been energetically unreachable for earlier hominins.
The stride length difference captures this in a single metric. Lucy’s estimated stride length was approximately 0.72 to 0.85 meters. Turkana Boy’s, even as a juvenile, was in the range of 1.1 to 1.5 meters — approaching the modern human average of 1.4 to 1.6 meters. Each stride covered more ground; each stride cost less energy. The compounding effect across a day of walking is enormous. This is the body that made the Out-of-Africa dispersal — documented at Dmanisi in Georgia by 1.77 million years ago — biomechanically and metabolically feasible. The legs came first. The continent followed.
Locomotion Biomechanics
Data: Pontzer 2012; Ruff & Walker 1993; Jungers 1988“Longer legs and a more efficient gait reduced the cost of transport in early Homo by roughly fifty percent compared to australopiths. That efficiency is what made long-distance ranging — and eventually continental dispersal — metabolically feasible.”
— Herman Pontzer, Current Anthropology, 2012
The Brain
“The brain was getting bigger, but it wasn’t yet the engine of change.”
The endocranial volume of KNM-WT 15000 has been measured at approximately 880 cubic centimeters. Given that hominin brain growth is roughly 95 to 97 percent complete by the equivalent of age seven to eight in modern humans, the projected adult cranial capacity falls in the range of 900 to 910 cc — substantially larger than any australopith (Lucy’s species averaged around 438 cc), larger thanHomo habilis (averaging approximately 610 cc), but well below the volumes recorded for later Homo erectus specimens from Asia (up to 1,100 cc) and dramatically below the modern human mean of approximately 1,400 cc.
It is tempting to place this number on a progress chart — to draw a line from 438 to 880 to 1,400 and call it the march of encephalization. But that line obscures more than it reveals. The brain of Homo ergaster was growing, yes, but the body was growing faster. Turkana Boy’s postcranial skeleton had modernized far more dramatically than his endocranium. His limb proportions, his pelvic geometry, his thermoregulatory architecture — all were within or approaching the modern human range. His brain was not. The encephalization quotient — brain size relative to expected brain size for a mammal of equivalent body mass — was elevated compared to australopiths but remained substantially below the modern human value. This is a body story first and a brain story second.
The range of endocranial volumes across Homo erectus sensu lato is itself instructive. Susan Antón, in her comprehensive 2003 review, documented a span from approximately 600 cc at Dmanisi to over 1,100 cc in later Asian specimens. Turkana Boy sits comfortably in the middle of this enormous range — a highly variable species in which brain size alone cannot distinguish populations, let alone predict behavioral capacity. The Dmanisi hominins, with their smaller brains, had already dispersed out of Africa. Larger brains did not appear to be a prerequisite for continental-scale movement.
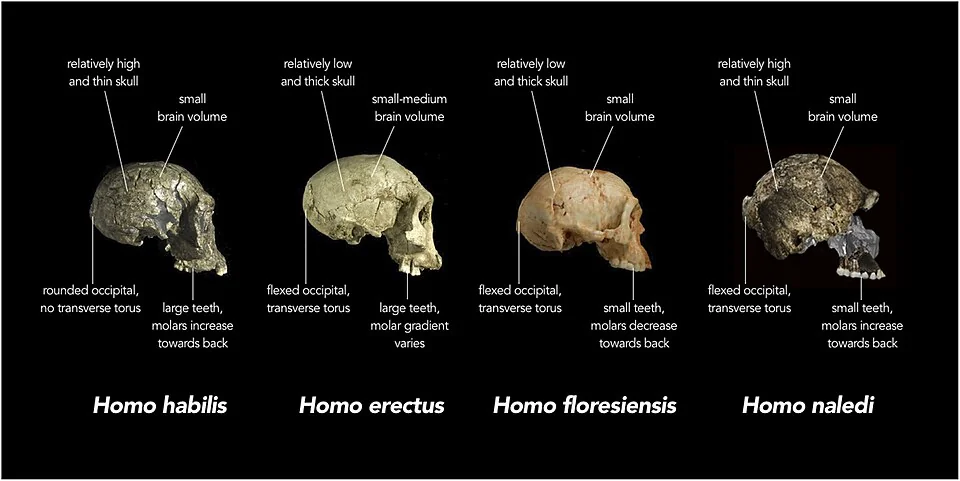
The most provocative cross-reference comes from a species that postdates Turkana Boy by over a million years. Homo naledi, dated to approximately 236,000 to 335,000 years ago in the Rising Star cave system of South Africa, had an endocranial volume of only 465 to 560 cc — smaller than many australopiths, far smaller than Turkana Boy, and yet the species was found in a context that suggests deliberate body disposal in a difficult-to-access underground chamber. If the mortuary behavior interpretation survives scrutiny, it means that a hominin with a brain smaller than Turkana Boy’s was engaged in symbolic or proto-symbolic activity nearly a million years later. Brain size alone does not determine cognitive capacity. Organization, connectivity, and developmental context all matter — and none of these can be read from a cranial volume measurement.

Endocranial Volume Across Hominin Species
Data: Holloway et al. 2004; Walker & Leakey 1993; Antón 2003“The range of endocranial volumes across Homo erectus sensu lato is enormous — from about 600 cubic centimeters at Dmanisi to over 1,100 in later Asian specimens. Turkana Boy at 880 sits comfortably in the middle of a highly variable species.”
— Susan Antón, Yearbook of Physical Anthropology, 2003
The Voice
“The question is not whether he could speak. The question is what the skeleton can and cannot tell us about it.”
There is a narrow canal running through the thoracic vertebrae of KNM-WT 15000 — the vertebral canal, the bony conduit that houses the spinal cord and its branching nerves. In the thoracic region, spinal nerves exit through the intervertebral foramina to become the intercostal nerves, which innervate the muscles between the ribs. These intercostal muscles do something that appears simple but is biomechanically extraordinary: they modulate exhalation. Not the gross inflation and deflation of breathing at rest, but the fine, graded control of airflow that allows a human being to sustain a vowel, vary the volume of a consonant, hold a phrase across a controlled breath, and — in the most complex manifestation — produce spoken language.
In 1999, Ann MacLarnon and Gwen Hewitt published a landmark analysis of the thoracic vertebral canal in KNM-WT 15000. Their finding was striking: the canal was significantly narrower than in modern humans, particularly in the upper and mid-thoracic regions where intercostal innervation is densest. MacLarnon and Hewitt argued that a narrower canal implied fewer nerve fibers passing through it, which in turn implied reduced motor innervation of the intercostal muscles. If the intercostal muscles received less neural input, they could not achieve the fine motor control of exhalation necessary for the sustained, modulated airflow that speech requires. The conclusion was carefully stated but widely cited: KNM-WT 15000 likely lacked the respiratory infrastructure for complex spoken language. The body was modern; the voice was not yet there.
The argument was elegant, anatomically grounded, and — as subsequent work would demonstrate — potentially flawed. In 2015, Marc Meyer and Martin Haeusler re-examined the vertebral canal dimensions using a substantially larger comparative sample of modern humans and fossil hominins. Their central finding challenged MacLarnon and Hewitt on two fronts. First, when measured against a broader range of modern human variation, the canal dimensions of KNM-WT 15000 fell within the lower end of the normal modern range rather than below it. Second, and more fundamentally, Meyer and Haeusler drew attention to the pathological condition of several of Turkana Boy’s vertebrae. Bruce Latimer and James Ohman had documented in 2001 that KNM-WT 15000 suffered from a form of axial dysplasia — a developmental abnormality of the vertebral column that affected vertebral body shape and canal dimensions. If pathology had distorted the canal, then measurements taken from the specimen could not be treated as representative of the species.
The disagreement between these two analyses is not merely technical. It illustrates a principle that governs much of paleoanthropology: the distance between a measurement and an inference is often greater than it appears. MacLarnon and Hewitt measured the canal. They inferred nerve count from canal width. They inferred motor control from nerve count. They inferred speech capacity from motor control. Each step is plausible; the chain is long. Meyer and Haeusler did not need to refute the logic — they only needed to question the first measurement, and the entire chain loosened. This is how science works when the sample size is one. A single specimen carrying a pathological condition can generate an inference about an entire species that survives for sixteen years before the measurement itself is reconsidered.
Neither analysis settles the question. MacLarnon and Hewitt may be correct thatHomo ergaster lacked the respiratory control for fully modern speech, even if their specific measurement was compromised by pathology — other evidence, including the relatively small size of the hypoglossal canal and the apparently high position of the larynx inferred from basicranial morphology, points in the same direction. Meyer and Haeusler may be correct that the vertebral canal alone cannot bear the inferential weight placed upon it. The honest summary is that speech in earlyHomo remains an open question, constrained by the limits of what bone can tell us about soft tissue, neural wiring, and behavior.
What the debate teaches, regardless of its resolution, is something about the nature of skeletal evidence. A vertebral canal is a channel for nerves. Nerves control muscles. Muscles produce movement, including the movement of air through a vocal tract. But the canal does not contain the nerves; they decomposed 1.53 million years ago. What remains is the housing, and from the housing we infer the contents, and from the contents we infer the capacity, and from the capacity we infer the behavior. At each step, certainty diminishes. The skeleton speaks — but in whispers, not declarations.
The Vertebral Canal Debate
MacLarnon & Hewitt 1999 vs. Meyer & Haeusler 2015MacLarnon & Hewitt (1999)
- Thoracic vertebral canal narrower than modern humans
- Reduced innervation of intercostal muscles
- Limited fine motor control of exhalation
- Complex speech requires precise breathing control
- Conclusion: speech capabilities were limited
Meyer & Haeusler (2015)
- Re-examined vertebral measurements with larger sample
- Canal width falls within modern human variation
- Pathological vertebrae may have distorted original analysis
- KNM-WT 15000 had vertebral anomalies (Latimer & Ohman 2001)
- Conclusion: original inference may overreach
“The thoracic vertebral canal of KNM-WT 15000 is significantly smaller than that of modern Homo sapiens. This implies reduced innervation of the intercostal muscles and, consequently, a more limited capacity for the fine respiratory control associated with spoken language.”
— Ann MacLarnon & Gwen Hewitt, American Journal of Physical Anthropology, 1999
The Tools
“A handaxe is a theory of shape held in the hand.”
In 2011, Christopher Lepre and colleagues published a finding that reset the chronology of stone tool technology. At the site of Kokiselei, on the western shore of Lake Turkana — roughly 50 kilometers from the spot where Kamoya Kimeu would find KNM-WT 15000 — they recovered a collection of large bifacially flaked tools from sediments dated by paleomagnetic methods to approximately 1.76 million years ago. These were not the simple, irregular flakes of the Oldowan tradition. They were Acheulean handaxes — large, symmetrically shaped, bifacially worked stones that require a fundamentally different manufacturing process from anything that preceded them. The Kokiselei discovery pushed the origin of the Acheulean back by roughly 350,000 years, placing the earliest known handaxes in the West Turkana Basin at a date contemporary with the Dmanisi hominins in Georgia and roughly 230,000 years before KNM-WT 15000 walked the same landscape.
The distinction between Oldowan and Acheulean technology is not merely typological — it is cognitive. Oldowan tools, first described from Olduvai Gorge in Tanzania and now documented as far back as 2.6 million years ago at Gona in Ethiopia, are produced by striking a hammerstone against a core to detach sharp-edged flakes. The process requires spatial coordination and an understanding of conchoidal fracture, but the resulting tools are irregular and show limited evidence of a predetermined form. The toolmaker is exploiting the physics of stone fracture; the shape emerges from the material rather than from a plan.
Acheulean handaxes are different. A finished handaxe displays bilateral symmetry — the tool is shaped to a template that exists, in some form, before the knapping begins. The toolmaker must rotate the core, remove flakes from alternating faces in a planned sequence, maintain proportionality between length and width, and correct asymmetries as they emerge. Dietrich Stout and colleagues, in a series of neuroimaging studies conducted between 2008 and 2015, demonstrated that Acheulean handaxe production activates prefrontal cortical regions associated with hierarchical action planning — regions that are not significantly activated during Oldowan flaking. The transition from Oldowan to Acheulean, in other words, tracks an increase not merely in manual skill but in the cognitive architecture supporting planned, multi-step, goal-directed behavior.

It would be tempting to place a handaxe in Turkana Boy’s hand. The Acheulean tradition was well established in West Turkana by 1.53 million years ago; Homo ergaster is the most likely maker; KNM-WT 15000 lived in the right place at the right time. But the temptation should be resisted. No tools were found in direct association with the skeleton. The Nariokotome site yielded the individual but not his toolkit. To assign specific technological competence to a specific fossil individual is to cross from population-level inference to individual biography, and the evidence does not support that crossing. What we can say is that the species to which KNM-WT 15000 belonged was the first to produce artifacts requiring a mental template — that the cognitive world his species inhabited included the capacity to impose planned, symmetrical form on raw stone. The handaxe is a theory of shape held in the hand. By 1.76 million years ago, that theory existed in West Turkana.


The broader technological context stretches both deeper into the past and further across the continent. At Lomekwi 3, also in West Turkana, Sonia Harmand and colleagues announced in 2015 the discovery of stone tools dated to 3.3 million years ago — the Lomekwian industry, predating even the Oldowan and potentially associated with a pre-Homo toolmaker. At Konso-Gardula in Ethiopia, Yonas Beyene and colleagues documented Acheulean handaxes at 1.75 million years ago, independently confirming the early emergence of the tradition in East Africa. The Acheulean would persist, with remarkable conservatism, for over a million years — the longest-lived technological tradition in human history, not superseded until the Middle Stone Age industries appeared after roughly 300,000 years ago. Turkana Boy lived at the front end of this extraordinary cultural persistence, in a landscape where the cognitive revolution encoded in bilateral symmetry was newly underway.
Stone Tool Technology Timeline
Data: Lepre et al. 2011; Harmand et al. 2015; Beyene et al. 2013“The Acheulian has now been shown to extend to 1.76 million years ago in West Turkana. This is substantially earlier than previously documented and indicates that the transition to large cutting tool technology was underway soon after the emergence of Homo erectus in East Africa.”
— Christopher Lepre et al., Nature, 2011
The Walk
“They did not need boats. They needed legs and heat tolerance and time.”
In 1991, a team led by Leo Gabunia and Abesalom Vekua recovered a mandible from the medieval ruins of Dmanisi in the Republic of Georgia. Within a decade, the site would yield five hominin crania and an extraordinary collection of postcranial remains, all dating to approximately 1.77 million years ago. Dmanisi is not in Africa. It is in the southern Caucasus, at the crossroads of Europe and Asia, perched above the confluence of the Mashavera and Pinezauri rivers at an elevation of roughly 800 meters. Its hominins — classified as early Homo erectus or Homo georgicus depending on the taxonomist — represent the earliest confirmed presence of our genus outside the African continent. They arrived roughly 240,000 years before KNM-WT 15000 was born.
The postcranial material from Dmanisi, published by David Lordkipanidze and colleagues in 2007, provides the critical comparison. The D4167 partial skeleton — comprising vertebrae, clavicle, humerus, ulna, portions of the os coxa, and a nearly complete tibia — displays body proportions that fall within the range documented for AfricanHomo ergaster. Estimated stature ranged from 145 to 166 centimeters depending on the regression method applied to the tibial length. Limb proportions were modern in their fundamental geometry: relatively long legs, relatively short arms, a linear body plan consistent with open-country bipedalism and efficient thermoregulation. The Dmanisi hominins were not identical to Turkana Boy — their brains were smaller (the smallest cranium, D4500, has an endocranial volume of roughly 546 cc, the largest about 730 cc), and their stature may have been somewhat shorter on average — but they shared the same fundamental body architecture. The engineering solution documented in KNM-WT 15000’s skeleton was already in place, in functional form, a quarter-million years earlier and 5,000 kilometers to the northeast.

The mechanism of dispersal does not require extraordinary explanation. It requires only legs, heat tolerance, and generational time. A home range expansion of 20 kilometers per generation — modest by the standards of large-bodied mammalian carnivores — would carry a population from East Africa to the Caucasus in roughly 25,000 years, a span that is archaeologically invisible and biologically trivial. The body plan documented in KNM-WT 15000 — the long stride, the efficient cost of transport, the thermoregulatory architecture optimized for sustained movement in hot, open environments — was the infrastructure of this dispersal. The hominins did not decide to leave Africa. They expanded their ranges in response to ecological opportunity, and their ranges eventually extended beyond the continent. The dispersal was not an event; it was a gradient.
Further evidence of early Homo outside Africa comes from the site of 'Ubeidiya in the Jordan Valley of Israel, where Acheulean tools and fragmentary hominin remains date to approximately 1.4 million years ago. The tools at 'Ubeidiya share typological characteristics with East African Acheulean assemblages, suggesting that the technological tradition traveled with the populations. By 1.0 million years ago, Homo erectus populations are documented across a vast swath of the Old World — from Java to northern China to southern Europe. The body plan that first appears clearly in the fossil record with specimens like KNM-WT 15000 proved to be one of the most successful anatomical designs in mammalian evolution, persisting with only modest variation for more than a million years across three continents.
The Dmanisi connection is particularly illuminating because it decouples brain size from dispersal ability. The Dmanisi hominins had cranial capacities well below Turkana Boy’s — yet they had already completed the journey. Whatever cognitive threshold continental dispersal requires, it had been met at 600 cc. The body, not the brain, was the primary enabling factor. Long legs that reduced the cost of transport. A narrow pelvis and linear build that shed heat across equatorial and subtropical landscapes. A stride length that could cover 30 to 40 kilometers in a day without extraordinary metabolic expenditure. These are not cultural innovations. They are anatomical facts, written in bone and readable in KNM-WT 15000, that made the first great human migration not just possible but, in hindsight, predictable.
Early Homo Dispersal: Key Sites
Data: Lordkipanidze et al. 2007; Lepre et al. 2011; Bar-Yosef & Goren-Inbar 1993“The Dmanisi postcranial evidence demonstrates that body proportions associated with modern-type locomotion and thermoregulation were established in early Homo well before the major increase in brain size seen in laterHomo erectus. The body came first.”
— David Lordkipanidze et al., Nature, 2007
The Finder
“He just picked it up. A fragment smaller than a matchbox. That was all it took.”
On Sunday morning, August 23, 1984, Kamoya Kimeu walked along a dry erosion gully near the Nariokotome River on the western shore of Lake Turkana. The area had been surveyed before. The team, led by Richard Leakey and Alan Walker, was prospecting for fossils in the Nachukui Formation — Plio-Pleistocene sediments exposed by the slow erosional retreat of ancient lake deposits. Kimeu was not looking for anything in particular. He was doing what he had done nearly every working day for over two decades: walking with his eyes on the ground, scanning the surface gravels for the distinctive texture and color of fossilized bone against a background of sedimentary debris. It is monotonous, physically demanding work that requires sustained attention over hours of equatorial heat. Most days yield nothing. Many weeks yield nothing. The entire enterprise depends on a form of expertise that resists formal description — an acquired perceptual skill, honed over thousands of hours, that allows certain individuals to detect the signal of bone in the noise of stone.
Kimeu spotted a small fragment of cranial bone lying on the slope of the gully. It was smaller than a matchbox — a piece of frontal bone, dark and mineralized, with the distinctive curvature and cortical thickness of a hominin cranium. He collected it and brought it to Leakey. The team returned to the spot. Within days, further surface collection and preliminary excavation revealed additional cranial fragments, and then a mandible, and then — astonishingly — postcranial elements: vertebrae, ribs, portions of the pelvis and limb bones. The scatter of bone extended across the hillslope in a pattern consistent with a single burial event followed by gradual erosional exposure. It became clear within the first field season that this was not a handful of fragments. This was a skeleton. An early Homo skeleton, the most complete one ever found by a factor that rendered comparisons meaningless.
Kamoya Kimeu is among the most accomplished fossil hunters in the history of paleoanthropology. Born near Kangundo in eastern Kenya around 1940, he began working with Louis Leakey in 1960 and went on to serve as field crew chief for both Richard Leakey and Meave Leakey across decades of expeditions in the Turkana Basin, at Olduvai Gorge, and at Laetoli. His discoveries include not only KNM-WT 15000 but also KNM-ER 1808 (a pathological Homo erectus femur from Koobi Fora), significant specimens of Homo habilis and Paranthropus boisei, and numerous other finds that collectively form a substantial fraction of the East African hominin fossil record. In 1985, the National Geographic Society awarded him the LaGorce Gold Medal — the first time the honor had been given for work in paleontology and only the second time it had been awarded to an African.
The story of discovery matters because it restores contingency to a process that textbooks often present as inevitable. KNM-WT 15000 was not found by remote sensing, geochemical survey, or algorithmic prediction. It was found because one person, on one morning, glanced at the right patch of ground. Had Kimeu walked ten meters to the left, the fragment might have eroded away in the next rainy season, washing into the Nariokotome River and eventually into Lake Turkana, where it would have been unrecoverable. The most consequential fossil find in the study of early Homo depended on an act of attention that was irreducible to method — a moment of perception by a trained eye in the right place at the right time.
This is not to diminish the systematic fieldwork that placed Kimeu at that location. Richard Leakey had selected the Nariokotome area based on geological mapping and prior reconnaissance. The team was working within a research framework that identified the Nachukui Formation as a promising fossiliferous horizon. But within that framework, the discovery itself was irreducibly human — an individual’s attention transforming an eroding scrap of bone into the pivot point of an entire scientific field. After eleven sections of anatomy, biomechanics, energetics, and cognitive inference, this is worth pausing to acknowledge: scientific revolutions sometimes begin not with hypotheses but with attention.
“Kamoya has a gift for finding things. I don’t know how he does it — none of us do, really. He just sees bone where the rest of us see rock. That morning he picked up a little scrap of cranium no bigger than a matchbox. That scrap turned out to be the most complete early human skeleton ever found.”
— Richard Leakey, Origins Reconsidered, 1992
The Excavation
“Five seasons. Fifteen hundred cubic meters. One skeleton.”
The discovery of a cranial fragment in August 1984 was the beginning, not the end, of the work. What followed was one of the most intensive and methodologically rigorous excavations in the history of paleoanthropology — five field seasons stretching from 1984 to 1989, during which the team led by Richard Leakey and Alan Walker systematically dismantled a hillslope to recover the remains of a single individual. The scale of the effort is difficult to convey without numbers: approximately 1,500 cubic meters of sediment were excavated and screened; every bucket of earth passed through wire mesh to capture fragments as small as tooth enamel chips; the excavation area eventually extended well beyond the initial scatter, following the geological logic of erosion and redeposition to locate elements that had migrated downslope.
The first season in 1984 established the scope of the find. The cranial fragments that Kimeu had spotted on the surface led the team to a concentration of bone eroding from a layer of fine-grained tuffaceous siltstone within the Nachukui Formation. By the end of the season, the team had recovered the cranium in multiple fragments, the mandible with its telltale abscess, and the first postcranial elements — cervical and thoracic vertebrae, several ribs, and portions of the upper limb. The realization that these all belonged to a single individual came from the congruence of size, developmental stage, and taphonomic context: every bone showed the same degree of mineralization, the same sedimentary matrix adhering to its surface, and the same juvenile developmental status.

The second and third seasons, in 1985 and 1986, yielded the bulk of the postcranial skeleton. Both femora were recovered largely intact — extraordinary finds given that long bones are among the most fragile elements in a juvenile skeleton, their unfused epiphyses vulnerable to post-mortem breakage and erosion. The tibiae, fibulae, and both innominate bones of the pelvis were recovered across these seasons, providing the critical material for the body proportion and locomotor analyses that would follow. The vertebral column was recovered in near-complete sequence: all twelve thoracic vertebrae, all five lumbar vertebrae, six of seven cervical vertebrae, and the sacrum. This degree of vertebral completeness is nearly unique in the hominin fossil record and made possible the vertebral canal analyses that would fuel the speech debate for the next two decades.
The fourth and fifth seasons, in 1987 and 1989, extended the excavation to its geological limits. Additional rib fragments, humeral pieces, and the few hand and foot elements that were recovered came from these later seasons. The final tally was 108 bones — representing approximately 40 percent of the complete skeleton. The missing elements followed a predictable taphonomic pattern: hands and feet, which are composed of numerous small bones loosely connected by soft tissue, are the most vulnerable to disarticulation and dispersal after death. The 54 bones of each hand and the 52 bones of each foot were almost entirely absent, likely scattered by scavengers or fluvial processes before burial was complete.
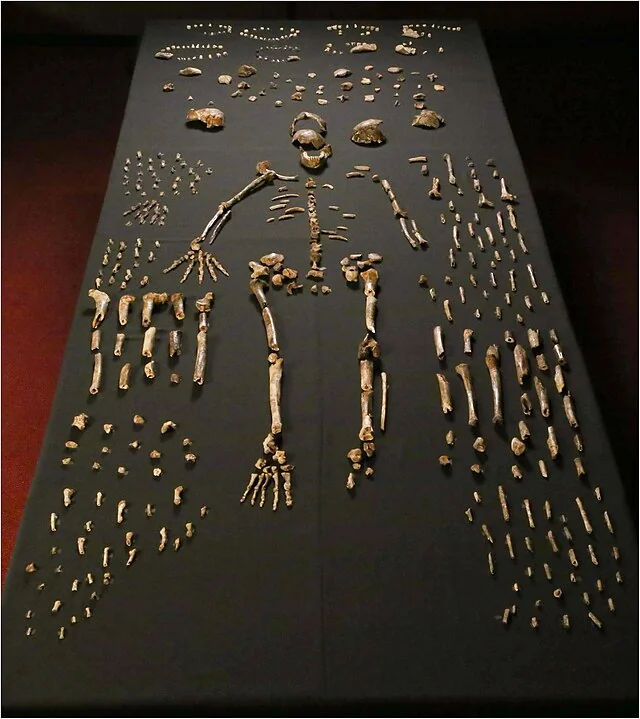
The resulting monograph — The Nariokotome Homo erectus Skeleton, edited by Walker and Leakey and published by Harvard University Press in 1993 — remains one of the most comprehensive analyses of a single fossil specimen ever produced. Its chapters, written by specialists across anatomy, developmental biology, paleontology, and geology, established the evidential baselines against which every subsequent study of KNM-WT 15000 has been measured. The monograph did not resolve every question — the age debate persists, the speech question remains open, the projected adult stature is still disputed — but it established the standard of documentation that makes resolution possible. It transformed a collection of bones into a permanent scientific resource, accessible to any researcher willing to engage the evidence on its own terms. This is how a skeleton becomes a benchmark: not through the drama of its discovery but through the discipline of its analysis.
Excavation Progress: Cumulative Bone Recovery (1984–1989)
Data: Walker & Leakey 1993“When all the bones were catalogued and laid out, what we had was unprecedented: 108 elements from a single juvenile Homo erectusindividual. Most hominin species are known from fragments — a jaw here, a femur there. This was a person. We could read his body the way you read a book — from top to bottom, system by system.”
— Alan Walker, The Nariokotome Homo erectus Skeleton, 1993
The Inheritance
“You are the living draft of an architecture that was drawn 1.5 million years ago.”
Look down at your own legs. The femur running from your hip to your knee is approximately 45 centimeters long in an average adult male, 43 in an average adult female. Turkana Boy’s was 43.2 centimeters — and he was a child with years of growth remaining, on even the most conservative model. Your tibia is proportionally long relative to your femur, giving you a crural index of approximately 83 to 85. His was 86. Your pelvis is narrow relative to your stature — a design optimized not for the wide, flaring stability of a body that still climbed trees, but for the efficient rotation of long legs beneath a vertically balanced trunk. His pelvis was narrower still. The architecture of your lower body — the long stride, the narrow waist, the high surface-area-to-mass ratio that keeps you from overheating on a summer run — was drafted 1.53 million years ago in a body that walked the western shore of Lake Turkana.
This is not a metaphor. It is a morphological claim, and a carefully bounded one. The body plan that first appears in the hominin fossil record with Homo ergaster — the linear, heat-adapted, long-legged, narrow-hipped design documented in KNM-WT 15000 — is, in its fundamental proportions, the body plan you are currently using to sit in a chair and read this essay. Your brain is three times larger. Your teeth are smaller. Your face is retracted beneath the frontal bone rather than projecting forward beneath a heavy brow ridge. But from the neck down, the distance between your body and Turkana Boy’s is far smaller than the distance between his body and Lucy’s. The postcranial revolution — the transition from a compact, arboreal-terrestrial compromise body to an elongated, open-country, thermoregulatory design — was largely complete by 1.5 million years ago. What followed, in the subsequent 1.5 million years, was primarily a cranial story: brain expansion, facial reduction, the development of a chin, the reshaping of the vocal tract. The body had already arrived at a design that would prove durable enough to carry our lineage across continents, through ice ages, and into the Holocene.
The claim is narrow but powerful: the ecological logic of the modern human body is already present in KNM-WT 15000. Not the full cognitive suite, not the symbolic capacity, not the language, not the social complexity — but the physical chassis. The legs that carry a modern marathon runner over 42 kilometers in two hours are operating the same biomechanical system that carried Homo ergaster across the savannas of the Turkana Basin. The thermoregulatory design that allows a human being to run in equatorial heat without fatal hyperthermia — the combination of eccrine sweating, low body hair, and high surface-area-to-mass ratio — was functional by 1.5 million years ago. We are running on inherited hardware.
The cross-references complete the picture. Lucy, at 3.18 million years ago, represents the ancestral condition: a committed biped, yes, but one still tethered to arboreal refuges, still carrying a compact body adapted to heat retention in a mosaic environment, still limited in ranging capacity by short legs and a high cost of transport. Homo naledi, at roughly 300,000 years ago, represents a cautionary footnote: a species with a tiny brain that may have engaged in complex behavior, reminding us that the relationship between anatomy and capability is never as simple as a progress chart implies. And Turkana Boy, at 1.53 million years ago, represents the pivot — the moment when the hominin body reached a configuration so effective that natural selection would largely conserve it, with only minor refinements, for the next 1.5 million years and counting. Your body is his body, modified but not revolutionized. The draft he carried is the draft you inhabit.
The dashboard below assembles the comparative data that has accumulated across the preceding sections — stature, mass, endocranial volume, crural index, femur length, humero-femoral index — for every species referenced in this essay. It is not a progress chart. It is a data table, and the honest reader will notice that progress is not what it shows. It shows variation, convergence, and the occasional surprise. It shows that the Dmanisi hominins reached Georgia with brains smaller than Turkana Boy’s. It shows that Homo heidelbergensis was heavier but not necessarily taller. It shows that the crural index — a measure of heat adaptation — peaked in KNM-WT 15000 and has not been exceeded since. The body we inherited is not the product of steady improvement. It is the product of ecological logic, applied once, decisively, at the boundary between the Pliocene world and the Pleistocene.
Comparative Dashboard: Hominin Body Plan Data
Data compiled from Walker & Leakey 1993; Ruff & Walker 1993; Lordkipanidze et al. 2007; Antón 2003; Holloway et al. 2004; Berger et al. 2015| Species | Stature (cm) | Mass (kg) | ECV (cc) | Crural Index | Femur (cm) | HF Index |
|---|---|---|---|---|---|---|
| A. afarensis | 107 | 29 | 438 | 84 | 28.0 | 84.6 |
| A. africanus | 126 | 35 | 450 | — | — | — |
| H. habilis | 118 | 34 | 610 | — | — | — |
| KNM-WT 15000 | 160 | 48 | 880 | 86 | 43.2 | 74 |
| H. erectus (Dmanisi) | 155 | 45 | 665 | 83 | 39.5 | — |
| H. erectus (later) | 172 | 58 | 950 | 84 | 45.5 | — |
| H. heidelbergensis | 172 | 70 | 1250 | 82 | 46.0 | 72 |
| H. sapiens | 171 | 70 | 1400 | 84 | 45.5 | 72 |
Cluster Context: Lucy — Turkana Boy — Homo naledi
Relative scoring (0–10) across five comparative axesHomo erectus Specimen Cabinet
3D scans · African Fossils / National Museums of Kenya, CC BY-NC-SAThere is a temptation, at the end of an essay like this, to reach for triumph. To declare Turkana Boy the ancestor, the prototype, the blueprint. But the evidence does not support triumphalism any more than it supports a simple progress narrative. KNM-WT 15000 was not your ancestor — not in any direct genealogical sense. He was a child who died of infection on the shore of a Pleistocene lake, and his lineage may well have ended without descendants. What he was, demonstrably and without interpretive overreach, is a member of the species that first assembled the body plan you currently inhabit. Not the mind, not the culture, not the language — the body. The long legs, the narrow waist, the heat-shedding surface, the stride that can cross a continent in generations. That architecture was his, and it is yours, and the connection between those two facts is not metaphorical. It is skeletal, measurable, and written in the proportions of bone.
He was eight years old, or eleven, depending on which clock you read. He stood 160 centimeters tall. He had an infected jaw. He walked a landscape of open savanna and seasonal rivers under an equatorial sun, in a body built to dissipate heat across every square centimeter of its linear, long-limbed surface. His species made handaxes, though we cannot place one in his hand. His species may or may not have had language, and the skeleton cannot resolve the question. What the skeleton can tell us — what 108 bones recovered across five field seasons from a hillside near the Nariokotome River do tell us, with the weight of measurement behind every claim — is that the human body, in its essential ecological design, was already here. Already walking. Already adapted. Already, in the deepest biomechanical sense, us.
“The shift in body proportions from Australopithecus to early Homo is the single most dramatic morphological transition in the hominin postcranial record. By the time of KNM-WT 15000, the essential architecture of the modern human body was in place — not the brain, not the face, but the body. The legs, the pelvis, the thermoregulatory design. We are still walking on that blueprint.”
— Synthesis from Walker & Leakey 1993; Ruff 2010; Pontzer 2012; Holliday 2012
Sources
- Brown, F. et al. (1985). An early Hominid skeleton from west Lake Turkana, Kenya. Nature, 316, 788-792.
- Walker, A. & Leakey, R. (eds.) (1993). The Nariokotome Homo erectus Skeleton. Harvard University Press.
- Smith, B.H. (1993). The physiological age of KNM-WT 15000. In Walker & Leakey (eds.), The Nariokotome Homo erectus Skeleton, 195-220.
- Dean, M.C. & Smith, B.H. (2009). Growth and development of the Nariokotome youth, KNM-WT 15000. In Grine, F. et al. (eds.), The First Humans, 101-120.
- Ruff, C.B. & Burgess, M.L. (2015). How much did KNM-WT 15000 weigh? Journal of Human Evolution, 81, 1-10.
- Ruff, C.B. & Walker, A. (1993). Body size and body shape. In Walker & Leakey (eds.), The Nariokotome Homo erectus Skeleton, 234-265.
- Wheeler, P.E. (1991). The influence of bipedalism on the energy and water budgets of early hominids. Journal of Human Evolution, 21, 117-136.
- Wheeler, P.E. (1993). The influence of the loss of functional body hair on the water budgets of early hominids. Journal of Human Evolution, 24, 13-28.
- MacLarnon, A. & Hewitt, G. (1999). The evolution of human speech: The role of enhanced breathing control. American Journal of Physical Anthropology, 109, 341-363.
- Meyer, M.R. & Haeusler, M. (2015). Spinal cord evolution in early Homo. Journal of Human Evolution, 88, 43-53.
- Lepre, C.J. et al. (2011). An earlier origin for the Acheulian. Nature, 477, 82-85.
- Lordkipanidze, D. et al. (2007). Postcranial evidence from early Homo from Dmanisi, Georgia. Nature, 449, 305-310.
- Pontzer, H. (2012). Ecological energetics in early Homo. Current Anthropology, 53(S6), S346-S358.
- Antón, S.C. (2003). Natural history of Homo erectus. Yearbook of Physical Anthropology, 46, 126-170.
- Rightmire, G.P. (1998). Human evolution in the Middle Pleistocene: The role of Homo heidelbergensis. Evolutionary Anthropology, 6, 218-227.
- Holliday, T.W. (2012). Body size, body shape, and the circumscription of the genus Homo. Current Anthropology, 53(S6), S330-S345.
- Latimer, B. & Ohman, J.C. (2001). Axial dysplasia in Homo erectus. Journal of Human Evolution, 40, 12-28.
- Graves, R.R. et al. (2010). Just-so stories and the origin of ecogeographic body shape variation in Homo. Journal of Human Evolution, 58, 495-504.
- Beyene, Y. et al. (2013). The characteristics and chronology of the earliest Acheulean at Konso, Ethiopia. PNAS, 110(5), 1584-1591.
- Smithsonian National Museum of Natural History, Human Origins Program — Homo erectus interpretive resources.
Image Credits
- Turkana Boy reconstruction at Neanderthal Museum — Neanderthal Museum / Wikimedia Commons, CC BY-SA 4.0
- Homo erectus reconstruction, Natural History Museum London — Wikimedia Commons, CC BY-SA 4.0
- Homo ergaster skull diagram — Jose-Manuel Benito / Wikimedia Commons, CC BY-SA 3.0
- Homo ergaster skull replica, World Museum Liverpool — Johnbod / Wikimedia Commons, CC BY-SA 3.0
- Hominin tibia from Koobi Fora — Blumenschine et al. 2023, Scientific Reports / Wikimedia Commons, CC BY 4.0
- Lake Turkana landscape — Wikimedia Commons, CC BY-SA 2.0
- Turkwel River and Lake Turkana — Wikimedia Commons, CC BY-SA 2.0
- Lake Turkana geographic map — Nicolas Eynaud / Wikimedia Commons, CC BY-SA 3.0
- East African Rift Valley from ISS — NASA / ISS Expedition 30, public domain
- Acacia Kenya savannah landscape — Wikimedia Commons, CC BY-SA 3.0
- Richard Leakey — Wikimedia Commons, CC BY-SA 3.0
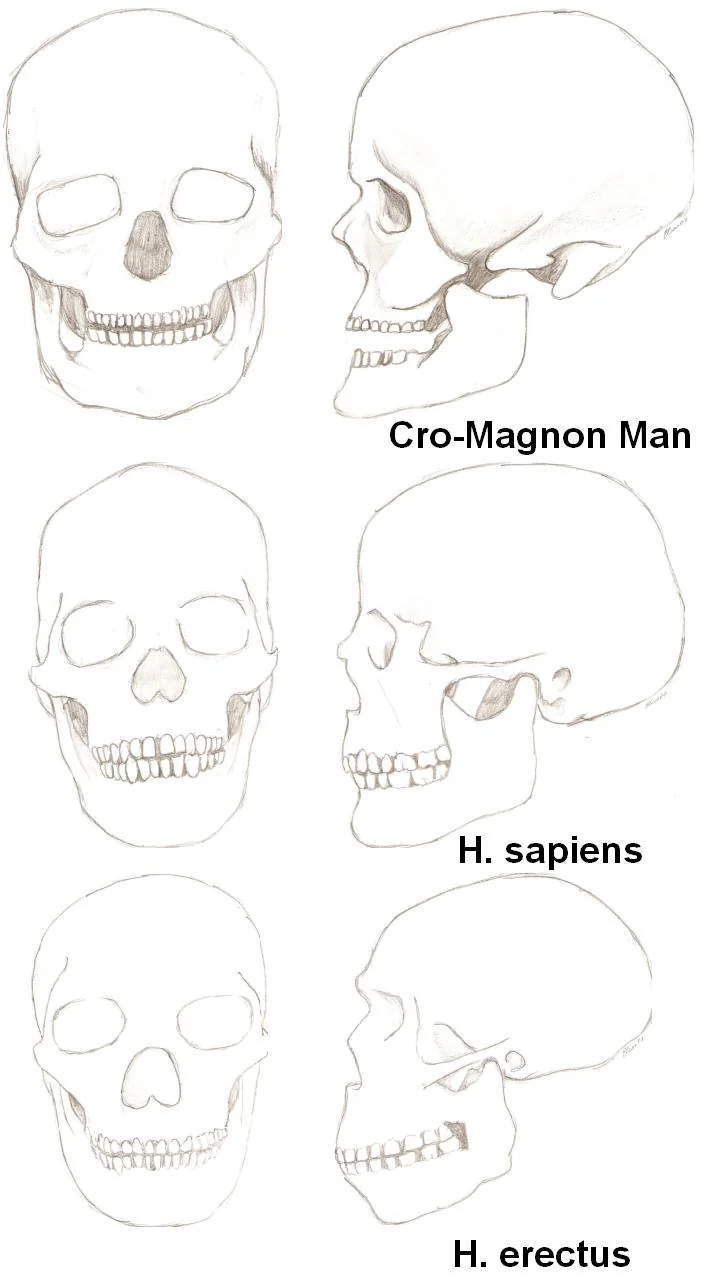
- Human evolution skeletal comparison (H. erectus, H. sapiens, Cro-Magnon) — Wikimedia Commons, CC BY-SA 3.0
- Comparison of skull features across early human species — Hawks et al. / Wits University / eLife, CC BY 4.0
- Dmanisi D3444 skull (Homo erectus / Homo georgicus) — Wikimedia Commons, CC BY-SA 3.0
- Lucy (A.L. 288-1) skeleton on black background — Museum national d'histoire naturelle / Wikimedia Commons, CC BY-SA 2.5
- Homo naledi skeletal specimens from Dinaledi Chamber — Berger et al. 2015, eLife, CC BY 4.0
- Acheulean bifaced ovate hand axe — Portable Antiquities Scheme / Wikimedia Commons, CC BY 2.0
- Bifaz de Atapuerca (Acheulean handaxe) — Wikimedia Commons, CC BY-SA 2.0
- Acheulean hand-axe from Egypt, Petrie Museum — Osama Shukir Muhammed Amin / Wikimedia Commons, CC BY-SA 4.0
- Homo erectus endocast, Smithsonian Museum — Tim Evanson / Wikimedia Commons, CC BY-SA 2.0
- Homo ergaster skull — Wikimedia Commons, CC BY-SA 4.0
- Homo ergaster skeleton (full view) — Wikimedia Commons, CC BY-SA 3.0