Homo naledi: The Small-Brained Species That Buried Its Dead
1,500 Bones, 560 Cubic Centimetres, and the Question of What Makes Us Human
The Recruitment
In October 2013, Lee Berger posted a message on Facebook. He was looking for scientists. Not senior researchers with decades of fieldwork experience, but small-bodied individuals willing to work in dangerous conditions deep underground, in spaces so confined that most adults could not physically enter them. The post was short, informal, and unprecedented in a discipline that had never recruited via social media. Within days, sixty applications arrived.
From that pool, Berger selected six women, all early-career researchers: Marina Elliott, Becca Peixotto, Alia Gurtov, Elen Feuerriegel, K. Lindsay Hunter, and Hannah Morris. He called them the “underground astronauts” — a name that captured both the physical extremity of their task and the strangeness of sending people into a space that no human being had entered in geological time. They would descend into the Rising Star cave system, navigate a 25-centimetre-wide passage that most adult males cannot fit through, climb over a jagged dolomite ridge called the Dragon’s Back, and lower themselves down a near-vertical chute into a chamber that had not seen natural light in at least hundreds of thousands of years.

Becca Peixotto and Marina Elliott during the Rising Star expedition. The six “underground astronauts” were recruited via social media — a first in paleoanthropology.
The paleoanthropology establishment reacted with skepticism. Berger had a complicated reputation: brilliant, media-savvy, polarizing. His discovery of Australopithecus sediba at Malapa in 2008 had been both celebrated and contested. The social-media recruitment struck traditionalists as a stunt. But the bones that the underground astronauts would bring to the surface over the next two years would force even the most skeptical critics to engage.
“The question was not whether we could get scientists down there. The question was whether we could get them back out.”— Lee Berger
The Cave
The Rising Star cave system lies within the Cradle of Humankind, a UNESCO World Heritage Site in Gauteng province, South Africa, roughly 50 kilometres northwest of Johannesburg. The cave opens in a dolomite hillside — limestone dissolved over millions of years by mildly acidic groundwater, creating a labyrinth of passages, chambers, and vertical shafts. It is this geology that makes the Cradle the richest single concentration of hominin fossil sites on Earth. Within a radius of 15 kilometres, the caves of Sterkfontein, Swartkrans, Kromdraai, Drimolen, and Malapa have yielded Australopithecus africanus,Paranthropus robustus, Australopithecus sediba, early Homo, and now Homo naledi.
The journey to the Dinaledi Chamber is a physical ordeal. From the cave entrance, cavers descend through Superman’s Crawl — a passage only 25 cm wide in places and12 m long, navigable only by individuals who can compress their bodies to fit through a slot narrower than a standard laptop. Beyond this lies the Dragon’s Back, a jagged ridge of exposed dolomite that must be climbed and then descended on the far side. Finally, a near-vertical chute drops 12 m into the Dinaledi Chamber itself — a confined space approximately 10 x 3 m with ceilings as low as one metre.

The Rising Star cave system during excavation. Live video feeds relayed from underground to a surface command tent allowed senior scientists to direct the work in real time.
Total darkness. No natural light has reached the Dinaledi Chamber in geological time. Every photograph, every observation, every millimetre of excavation was performed by headlamp. The underground astronauts worked in shifts, communicating with the surface via live video relay, receiving instructions from Berger and senior team members who could not physically access the chamber themselves. Specimens were passed hand to hand through the chute and along the narrow passages in padded bags — a logistics chain designed to move fossils without damaging material that had survived hundreds of thousands of years in darkness.
The Haul
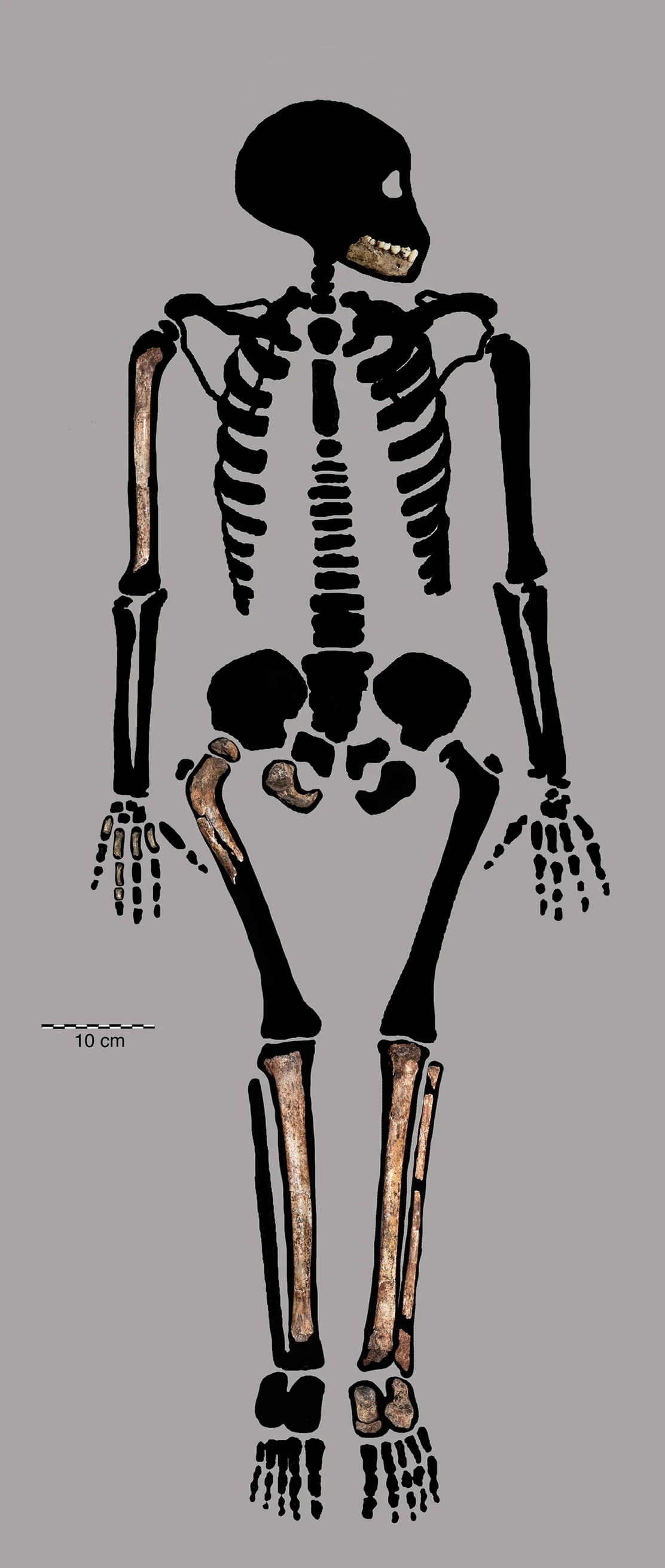
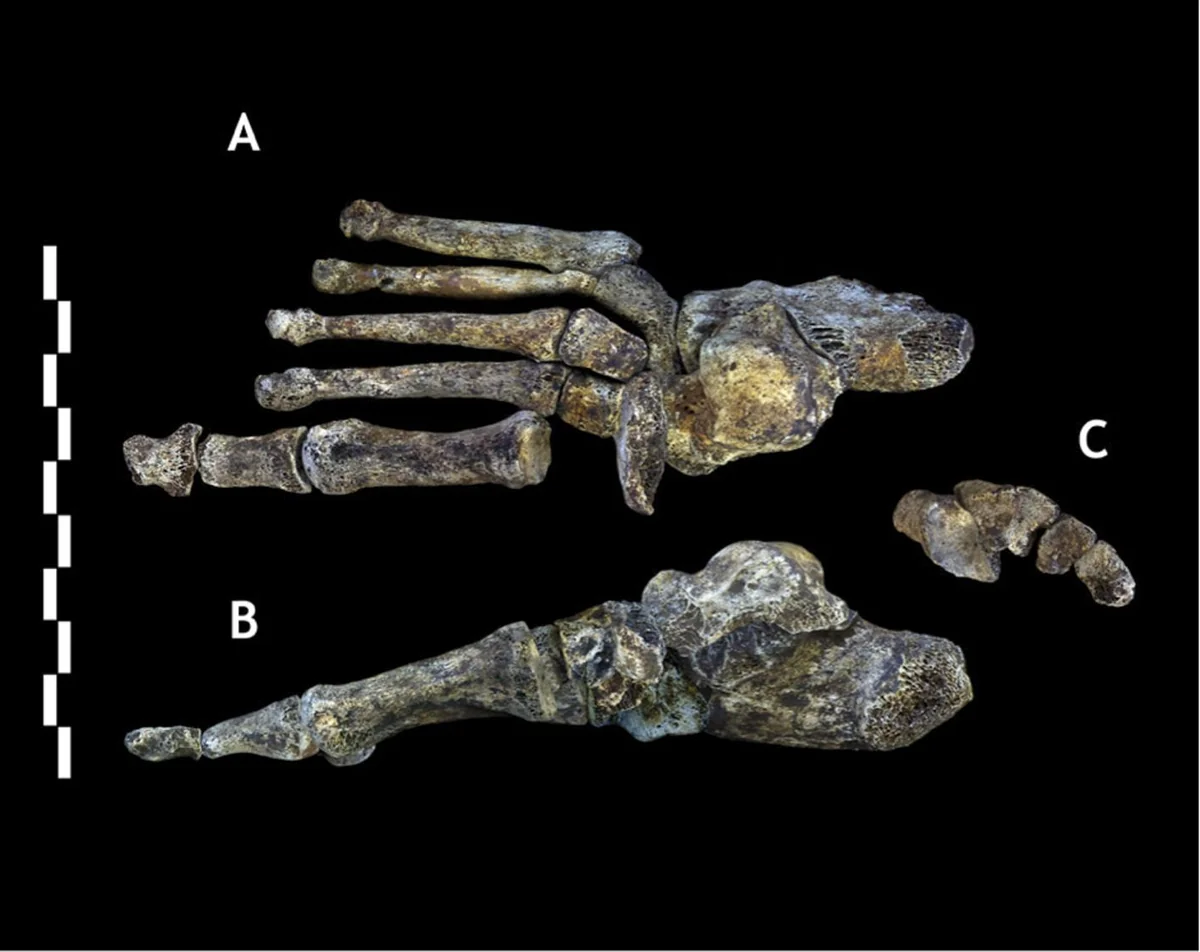
Over two field seasons, the underground astronauts recovered over 1,550 fossil specimens from the Dinaledi Chamber, representing at least 15 individuals spanning infants, juveniles, adults, and elderly. This made the Dinaledi assemblage one of the largest single-species hominin collections in the history of African paleoanthropology. For context: many hominin species are known from a single jawbone or partial skull. Homo rudolfensis was described from a single cranium.Kenyanthropus platyops rests on one distorted skull. Naledi offered an embarrassment of riches — crania, mandibles, vertebrae, ribs, pelves, long bones, hands, and feet, enough material to describe a species in extraordinary anatomical detail.
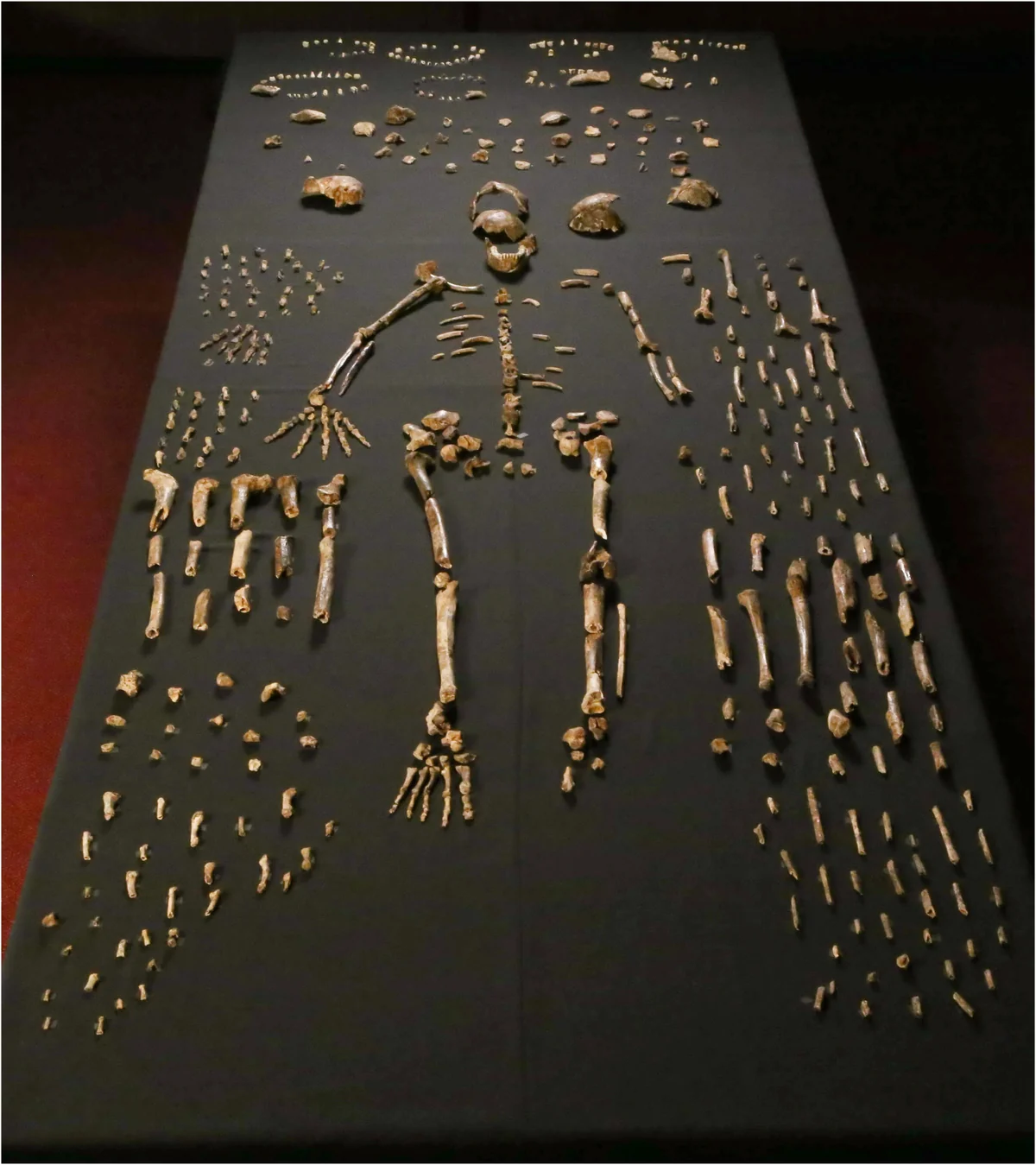
The Dinaledi skeletal assemblage. Over 1,550 fossil elements were recovered from at least 15 individuals — one of the largest single-species hominin collections in Africa.
The scale of the find was itself an anomaly. In most fossil sites, bones accumulate through a combination of predation, flooding, and chance — a leopard dragging a carcass into a cave, a river washing remains into a sinkhole. Such processes produce mixed assemblages: the bones of many species, scattered and fragmentary. The Dinaledi Chamber contained almost exclusively Homo naledi. No antelope. No predator remains. No evidence of the processes that normally explain how bones arrive in caves. This absence was the first hint that something unusual had happened in this chamber — a question that would dominate the scientific debate for the next decade.
The Mosaic Body
Homo naledi is not a transitional form. It is a mosaic — a species that combines anatomical features separated by millions of years in other lineages, assembled into a body plan that is stable, functional, and without clear precedent. The hands have curved proximal phalanges suited to tree-climbing alongside a long thumb and modern wrist morphology capable of precision grip and, potentially, tool manufacture. The feet are virtually modern — arched, with an adducted hallux, built for committed bipedal locomotion. The shoulders are primitive, oriented cranially in a configuration more similar to australopiths than to any species of Homo. The pelvis shows flared iliac blades reminiscent of Australopithecus. The teeth are small and simple, more Homo-like than australopith. And the brain — the brain is the crux of everything — is approximately 560 cc, within the range of a gorilla.
| Region | Classification | Details |
|---|---|---|
| Brain | primitive | ~560 cc (gorilla-range) |
| Brow ridges | primitive | Moderate supraorbital torus |
| Teeth | derived | Small, simple crowns (Homo-like) |
| Shoulders | primitive | Cranially oriented (australopith-like) |
| Trunk | primitive | Funnel-shaped thorax |
| Hands (fingers) | primitive | Curved proximal phalanges |
| Hands (thumb/wrist) | derived | Long thumb, modern wrist |
| Pelvis | primitive | Flared iliac blades |
| Legs | derived | Long, modern proportions |
| Feet | derived | Arch, adducted hallux (fully modern) |
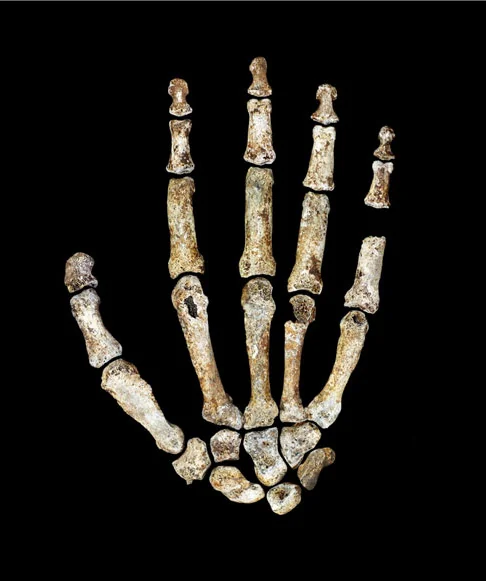
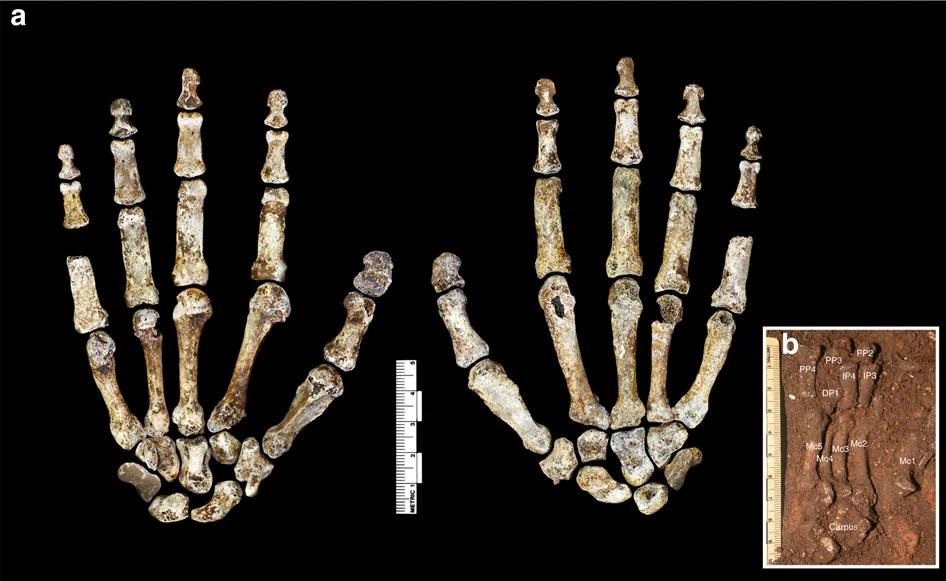
The combination is significant because it resists easy classification. Naledi is not an early Homo on its way to becoming something more modern. It is not a late australopith clinging to ancestral features. It is a distinct configuration — a species that assembled traits from different evolutionary eras into a body plan that persisted, successfully, for an unknown but potentially very long period. As John Hawks observed: “It’s as if evolution assembled this creature from a parts bin spanning three million years.”
The Brain Problem
560 cc. The average endocranial volume of Homo naledi, based on measurements from multiple specimens (range: approximately 465--610 cc). For context: a modern gorilla averages about 500 cc. Homo erectus, the species most commonly associated with the first major expansion of brain size in the genus Homo, averages approximately 900 cc. Modern humans average roughly1,400 cc. Naledi’s brain is one-third the size of ours, and smaller than most australopiths.
H. naledi appears twice: if deliberate disposal is accepted (score 7) or rejected (score 3). The species is an outlier either way — its brain size predicts far less behavioural complexity than it may have exhibited.
But endocranial volume is not the whole story. Holloway and colleagues (2018) examined virtual endocasts of naledi and found that despite the small volume, the frontal lobe organization resembles that of Homorather than australopiths. The brain is not merely a scaled-down ape brain — it shows structural reorganization in regions associated with planning, social cognition, and possibly language precursors. This does not prove that naledi was capable of complex behaviour. But it suggests that volume alone is not the right metric for assessing cognitive potential.
The Dating Bombshell
When Homo naledi was announced in 2015, no dates were published. The primitive morphology led most specialists to assume the species was ancient — perhaps 2 million years old or more, contemporary with early Homo habilis or late australopiths. The small brain, the australopith-like shoulders and pelvis, the curved fingers — everything about the anatomy suggested deep time.
In 2017, Dirks and colleagues published the results of three independent dating methods applied to the Dinaledi Chamber deposits. Uranium-thorium dating of flowstones bracketing the fossils. Electron spin resonance (ESR) dating of three naledi teeth. Palaeomagnetic analysis of the sediment sequence. All three methods converged on a single, startling answer: 236,000--335,000 years ago. Not two million years. Not even one million. Naledi was a contemporary of archaic Homo sapiens.
The implications were immediate and profound. A species with a brain one-third the size of ours, with a body plan mixing features from vastly different evolutionary eras, was alive in southern Africa at the same time as the earliest members of our own species. The landscape of the mid-Pleistocene was not a simple progression from primitive to modern. It was a mosaic of coexisting lineages — big-brained and small-brained, archaic and derived, living side by side in a small patch of southern African dolomite.
“When the dates came back, nobody believed them. We ran them again. Same answer.”— Paul Dirks
The Disposal Debate
The central question of the naledi story is not anatomical but behavioural: how did over 1,500 bones from at least 15 individuals end up in a chamber that is nearly impossible to reach? The taphonomic evidence — the study of what happens to organisms after death — eliminates most conventional explanations. No predator tooth marks. No water-transported sediment. No other large fauna. No evidence of a mass death event. No alternative entrance. What remains, by elimination, is the hypothesis that the bodies were placed there deliberately.
The word “disposal” matters. Berger’s team is careful to distinguish deliberate body disposal from burial. Burial implies grave-digging, grave goods, ritual intent — behaviours associated exclusively with large-brained species. Disposal means only that bodies were intentionally carried to this location and deposited. The cave itself is the container. No grave was dug. No objects were placed with the remains. The claim is modest in its specifics but revolutionary in its implications: a species with a 560 cc brain may have engaged in a community-level practice of dealing with its dead.
The counter-arguments are serious. Thackeray (2016) proposed that a second entrance may have existed and subsequently collapsed, eliminating the need for a deliberate journey through the existing passages. Others have suggested death-trap scenarios: animals falling in one at a time over millennia. The representation of all age classes (infants through elderly) is consistent with deliberate disposal but could also result from a long-duration death trap. The scientific community remains divided, and the debate is far from settled.
“We are not claiming ritual. We are claiming that these bodies were placed here. The cave itself is the evidence.”— Lee Berger
Neo and Lesedi
In 2017, Hawks and colleagues announced the discovery of a second deposit of Homo naledi remains in the Lesedi Chamber — a separate part of the Rising Star cave system, accessed by a different (equally difficult) route. The Lesedi assemblage included the most complete naledi skull ever found, nicknamed “Neo” (Sesotho for “a gift”), along with postcranial elements from at least three individuals.
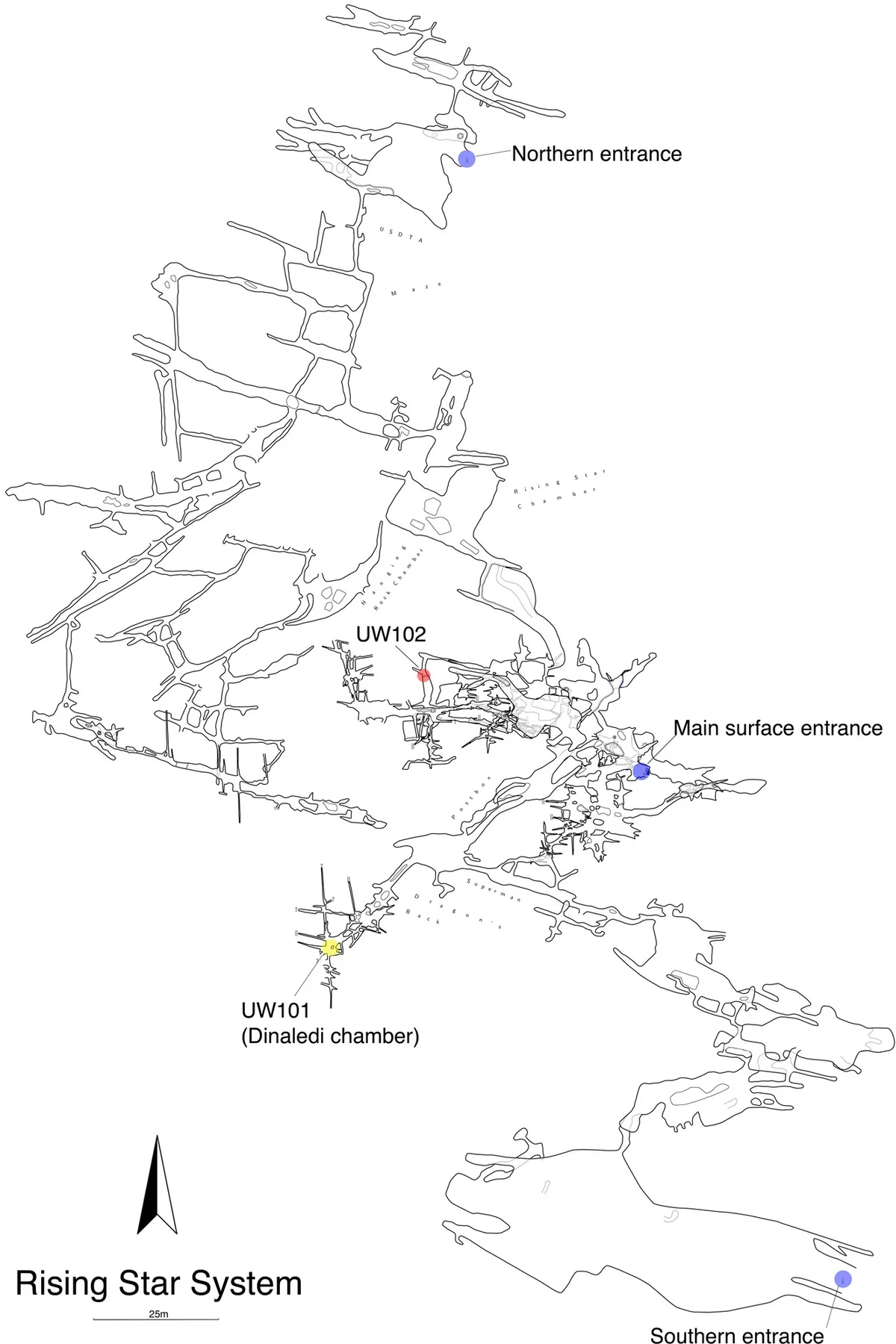
The Rising Star cave system, showing both the Dinaledi and Lesedi Chambers. Two independent deposits of H. naledi in isolated chambers strengthens the deliberate disposal hypothesis.
Same species. Different chamber. Same apparent depositional pattern: bodies in an isolated, nearly inaccessible space with no other large fauna. The discovery of a second deposit was significant precisely because it demonstrated a pattern. One chamber might be explained by unusual circumstances. Two chambers, in different parts of the same cave system, both containing only naledi remains, both accessed only through extreme physical difficulty — that looks less like accident and more like intent.
“Finding them once is extraordinary. Finding them twice, in the same way, in a different part of the cave — that’s a pattern.”— Marina Elliott
Fire and Engravings
In 2023, Berger’s team announced two additional findings from the Rising Star system: evidence of controlled fire use (hearths, soot deposits on cave ceilings, charred bone fragments) and geometric markings on cave walls (cross-hatch patterns incised into dolomite surfaces). If confirmed, these claims would place Homo naledi among the very few species known to have used fire and produced symbolic markings — behaviours previously associated only with large-brained hominins.
The scientific community’s response was largely skeptical. The results were announced via preprint and a Netflix documentary before surviving full peer review, a sequence that drew sharp criticism from researchers who argued that extraordinary claims demand extraordinary scrutiny. Tim White, a prominent paleoanthropologist, noted that the evidence presented did not meet the threshold required to overturn established models of cognitive evolution. The hearth evidence, in particular, has been questioned on taphonomic grounds: soot deposits in caves can result from natural fires, and the spatial association between charred material and naledi remains has not been established to the satisfaction of most specialists.
What would need to be true for these claims to hold: if a 560 cc brain produced controlled fire, geometric marking, and deliberate body disposal, then virtually every existing model linking brain size to behavioural complexity would require fundamental revision. That threshold has not yet been crossed. The fire and engraving claims remain contested, and the scientific process continues.
The Open Science Revolution
Beyond the bones themselves, the naledi project disrupted the practice of paleoanthropology. Berger’s team published in eLife (open-access) rather than Nature or Science (paywalled), making all data freely available on the day of publication. Every specimen was digitized and the 3D surface scans released on MorphoSource under open-access educational licensing — meaning any researcher on Earth could study naledi without visiting South Africa, without requesting permission, without waiting years for access that might never come.

Museum reconstruction of Homo naledi at the Trento Science Museum (MUSE). Open-access 3D scans have enabled museums worldwide to produce accurate reconstructions.
The turnaround was historically fast. From initial excavation in 2013 to species description in 2015 — roughly two years. In a field where some fossil discoveries have taken decades to reach publication (theArdipithecus remains found in 1994 were not fully published until 2009), this pace was extraordinary and controversial. Traditionalists accused Berger of rushing to publish, of prioritizing media attention over scientific rigour. Supporters countered that open access and rapid publication served science better than decade-long embargoes that kept fossils locked in vaults.
“If you lock fossils in a vault, they belong to one scientist. If you put them online, they belong to the species.”— Lee Berger
Where Does Naledi Fit?
The phylogenetic position of Homo naledi remains unresolved. Bayesian analyses by Dembo and colleagues (2016) placed naledi in three plausible positions, none of which can be confidently excluded on current evidence. Model A positions naledi as basal Homo — branching off near the root of the genus, a very early experiment in the Homo body plan. Model B places naledi as sister toHomo erectus — sharing a recent common ancestor with the lineage that eventually led to us. Model C treats naledi as an independent long-surviving lineage with no close living relatives, evolving in parallel for potentially millions of years.

Gauteng province, South Africa. The Cradle of Humankind — home to Rising Star, Sterkfontein, Swartkrans, Malapa, and Drimolen — occupies an area smaller than the city of London.
The comparison with Australopithecus sediba is instructive. Sediba, also discovered by Berger (at Malapa, 2008), is another mosaic species that resists easy classification — dated to approximately 1.98 Ma, combining australopith and Homofeatures in proportions that have generated similar phylogenetic debate. Two mosaic species from the same researcher, in the same region, both defying neat categorization: pattern or coincidence? The honest answer is that we do not know. The South African cave record is revealing a hominin diversity that the traditional family tree cannot easily accommodate.
What the Small Brain Means
If 560 cc can carry its dead through pitch-dark passages and deposit them in remote chambers, then brain size is not the threshold for complex behaviour we assumed it was. This is the proposition that naledi forces upon us. It does not require accepting every claim Berger’s team has made — the fire and engraving evidence remains contested, the disposal hypothesis is debated, the phylogenetic position is unresolved. Even the most conservative reading of the evidence raises questions that have no comfortable answers.
The metric is wrong. Endocranial volume, treated for a century as the primary proxy for cognitive capacity in hominins, captures only one dimension of a multidimensional phenomenon. Brain organization, neural connectivity, life history, social complexity, ecological context — all of these shape what a species can do. Holloway’s endocast work on naledi shows a reorganized frontal lobe in a small braincase. The assumption that bigger always means smarter was convenient but may have been misleading.
The thread connects backward to Lucy. Australopithecus afarensis walked upright at438 cc — bipedalism did not require a big brain. Now naledi suggests that mortuary behaviour may not require one either. The “big brain first” narrative keeps failing. Each new discovery erodes the assumption that our brain is what made us special, that cognition was the engine and everything else followed.

Facial reconstruction of Homo naledi, produced using the coherent anatomical deformation technique. A face from 300,000 years ago — contemporary with the first members of our own species.
What does “human” mean if brain size is not the dividing line? The question naledi poses is not yet answered. The fire claims remain unresolved. The phylogenetic position is debated. New chambers in the Rising Star system may still hold discoveries that rewrite the story again. What is certain is that a small-brained species with a mosaic body lived in southern Africa at the same time as early Homo sapiens, and that explaining how 1,500 of its bones ended up in an unreachable chamber remains one of the most compelling puzzles in paleoanthropology.
“We have been telling ourselves a story about big brains for a hundred years. Naledi asks whether we were listening to the wrong story.”— Lee Berger
Sources & Further Reading
- 1.Berger, L.R. et al. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife, 4:e09560.
- 2.Dirks, P.H.G.M. et al. (2015). Geological and taphonomic context for the new hominin species Homo naledi from the Dinaledi Chamber, South Africa. eLife, 4:e09561.
- 3.Hawks, J. et al. (2017). New fossil remains of Homo naledi from the Lesedi Chamber, South Africa. eLife, 6:e24232.
- 4.Dirks, P.H.G.M. et al. (2017). The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife, 6:e24231.
- 5.Kivell, T.L. et al. (2015). The hand of Homo naledi. Nature Communications, 6:8431.
- 6.Harcourt-Smith, W.E.H. et al. (2015). The foot of Homo naledi. Nature Communications, 6:8432.
- 7.Garvin, H.M. et al. (2017). Body size, brain size, and sexual dimorphism in Homo naledi from the Dinaledi Chamber. Journal of Human Evolution, 111, 119–138.
- 8.Val, A. (2016). Deliberate body disposal by hominins in the Dinaledi Chamber, Cradle of Humankind, South Africa? Journal of Human Evolution, 96, 145–148.
- 9.Thackeray, J.F. (2016). The possibility of lichen growth on bones of Homo naledi: Were they exposed to light? South African Journal of Science, 112(7/8).
- 10.Berger, L.R. et al. (2023). Evidence for deliberate burial of the dead by Homo naledi. eLife, 12:RP89106.
- 11.Martinón-Torres, M. et al. (2015). Dental morphology of Homo naledi. Nature Communications, 6:8431.
- 12.Williams, S.A. et al. (2017). The vertebrae and ribs of Homo naledi. Journal of Human Evolution, 104, 136–154.
- 13.Dembo, M. et al. (2016). The evolutionary relationships and age of Homo naledi: An assessment using dated Bayesian phylogenetic methods. Journal of Human Evolution, 97, 17–26.
- 14.Elliott, M.C. et al. (2021). Underground astronauts: The excavation methodology of the Rising Star Cave system. South African Archaeological Bulletin, 76(215), 115–128.
- 15.Shreeve, J. (2015). Mystery of the New Human Species. National Geographic, 228(4), 30–57.
- 16.Holloway, R.L. et al. (2018). Endocast morphology of Homo naledi from the Dinaledi Chamber, South Africa. Proceedings of the National Academy of Sciences, 115(22), 5738–5743.
- 17.Feuerriegel, E.M. et al. (2017). The upper limb of Homo naledi. Journal of Human Evolution, 104, 155–173.
- 18.Marchi, D. et al. (2017). The thigh and leg of Homo naledi. Journal of Human Evolution, 104, 174–204.
- 19.VanSickle, C. et al. (2017). Homo naledi pelvic remains from the Dinaledi Chamber, South Africa. Journal of Human Evolution, 104, 185–206.
- 20.Laird, M.F. et al. (2017). The skull of Homo naledi. Journal of Human Evolution, 104, 100–123.
- 21.Schroeder, L. et al. (2017). Skull diversity in the Homo lineage and the relative position of Homo naledi. Journal of Human Evolution, 104, 124–135.
- 22.Cofran, Z. & Walker, C.S. (2017). Dental development in Homo naledi. Journal of Human Evolution, 104, 34–44.
- 23.Zipfel, B. et al. (2017). The foot and ankle of Homo naledi from the Dinaledi Chamber, South Africa. Journal of Human Evolution, 104, 155–173.
- 24.Wood, B. & Boyle, E.K. (2016). Hominin taxic diversity: Fact or fantasy? American Journal of Physical Anthropology, 159(S61), 37–78.
Image Credits
- Homo naledi skeletal specimens from Dinaledi Chamber — Berger et al. 2015, eLife (CC-BY 4.0)
- Homo naledi facial reconstruction (coherent anatomical deformation) — Cicero Moraes, CC-BY-SA 4.0
- Homo naledi facial reconstruction (MUSE museum) — Matteo De Stefano/MUSE, CC-BY-SA 3.0
- Rising Star cave exploration — Wits University/Berger, CC-BY 2.0
- Cross-section of Rising Star Cave system and Dinaledi Chamber — Paul H.G.M. Dirks et al. 2015, eLife (CC-BY 4.0)
- Rising Star Cave system showing Dinaledi and Lesedi Chambers — Hawks et al. 2017, eLife (CC-BY 4.0)
- Homo naledi right hand, dorsal view — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi Hand 1, complete adult right hand — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi Hand 1 wrist bones — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi first metacarpals — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi pollical metacarpal (Mc1) morphology — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Phalangeal curvature in Homo naledi — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi foot composite — Harcourt-Smith et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi adult right foot — Harcourt-Smith et al. 2015, Nature Communications (CC-BY 4.0)
- Digital reconstruction of Homo naledi foot — Harcourt-Smith et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi tali from Dinaledi Chamber — Harcourt-Smith et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi mandible shortly after recovery — Lee R. Berger, eLife (CC-BY 4.0)
- Mandibular and dental anatomy comparison across Homo species — Hawks et al. 2017, eLife (CC-BY 4.0)
- DH7 Homo naledi cranium — Berger et al. 2015, eLife (CC-BY 4.0)
- Homo naledi second metacarpal UW 101-1474 — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- Homo naledi museum reconstruction (Trento Science Museum) — Sailko, CC-BY 3.0
- Marina Elliott, underground astronaut and paleoanthropologist — Marina Elliott, CC-BY-SA 4.0
- Becca Peixotto, underground astronaut — Becca Peixotto, CC-BY-SA 4.0
- Becca Peixotto and Marina Elliott during Rising Star expedition — Wits University/Berger, CC-BY 2.0
- Mrs Ples (Sts 5) Australopithecus africanus skull — Didier Descouens, CC-BY-SA 4.0
- South Africa Gauteng province location map — NordNordWest, CC-BY-SA 3.0
- Homo naledi Hand 1 non-pollical metacarpals — Kivell et al. 2015, Nature Communications (CC-BY 4.0)
- 3D skull and mandible reconstruction (DH1/DH3 composite) — Peter Schmid/Berger et al. 2015, MorphoSource (Open Download), Rising Star Project
- 3D Hand-1 composite laser scan — Bolter/Berger et al. 2015, MorphoSource (Open Download), Rising Star Project
All images sourced from Wikimedia Commons under Creative Commons or Public Domain licenses. Specimen photographs from eLife publications (CC-BY 4.0). Anatomical and dating data drawn from cited primary literature.
Further Reading
- Seven Million Years: The Complete Visual History of Our Kind — The parent essay covering seven million years of hominin evolution, from Sahelanthropus tchadensis to Homo sapiens, with naledi’s place in the wider story.
- Turkana Boy: The Skeleton That Redefined Human Evolution — KNM-WT 15000, the most complete early human skeleton ever found. An interactive visual essay on the 1.5-million-year-old Homo erectus youth with 3D fossil specimens from the Turkana Basin.
- Lucy: Before the Genus Homo — A deep-dive visual essay on AL 288-1, the 3.2-million-year-old Australopithecus afarensis skeleton that rewrote the story of walking upright.
- Infographic: How Our Brains Grew Over 7 Million Years — Cranial capacity comparison across 8 hominid species, from Sahelanthropus (350 cc) to Homo sapiens (1,350 cc).